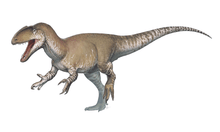Coelophysis rhodesiensis
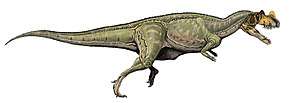
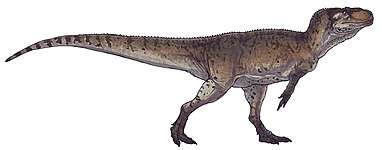
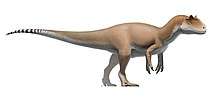
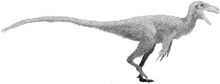
Coelophysis rhodesiensis is an extinct species of coelophysid theropod dinosaur that lived approximately 188 million years ago during the early part of the Jurassic Period in what is now Africa. The species was a small to medium-sized, lightly built, ground-dwelling, bipedal carnivore, that could grow up to 3 m (9.8 ft) long. It was formerly called Syntarsus, but that name was already taken by a beetle, and was subsequently given the name Megapnosaurus by Ivie, Ślipiński & Węgrzynowicz, in 2001, though many subsequent studies have classified it in the genus Coelophysis.
| Coelophysis rhodesiensis | |
|---|---|
 | |
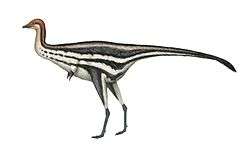
| Life restoration | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Family: | †Coelophysidae |
| Genus: | †Coelophysis |
| Species: | †C. rhodesiensis |
| Binomial name | |
| †Coelophysis rhodesiensis (Raath, 1969) | |
| Synonyms | |
| |
Description
Coelophysis rhodesiensis measured up to 3 meters (10 ft) long from nose to tail and weighed about 32 kilograms (70 lb). The bones of 30 C. rhodesiensis individuals were found together in a fossil bed in Zimbabwe, so paleontologists think it may have hunted in packs. The various fossils attributed to this species have been dated over a relatively large time span – the Hettangian, Sinemurian, and Pliensbachian stages of the Early Jurassic – meaning the fossils represent either a highly successful genus or a few closely related animals all currently assigned to Coelophysis.
Specimen UCMP V128659 was discovered in 1982 and referred to Megapnosaurus kayentakatae by Rowe (1989),[1] as a subadult gracile individual and later, Tykoski (2005)[2] agreed. Gay (2010) described the specimen as the new tetanurine taxon Kayentavenator elysiae,[3] but Mortimer (2010) pointed out that there was no published evidence that Kayentavenator is the same taxon as M. kayentakatae.[4]
Classification
|
"Syntarsus" rhodesiensis was first described by Raath (1969) and assigned to Podokesauridae.[6] The taxon "Podokesauridae", was abandoned since its type specimen was destroyed in a fire and can no longer be compared to new finds. Over the years paleontologists assigned this genus to Ceratosauridae (Welles, 1984), Procompsognathidae (Parrish and Carpenter, 1986) and Ceratosauria (Gauthier, 1986). Most recently, it has been assigned to Coelophysidae by Tykoski and Rowe (2004), Ezcurra and Novas (2007) and Ezcurra (2007), which is the current scientific consensus.[7]
According to Tykoski and Rowe (2004) Coelophysis rhodesiensis can be distinguished based on the following characteristics:[7] it differs from Coelophysis bauri in the pit at the base of the nasal process of the premaxilla; it differs from C.? kayentakatae because the promaxillary fenestra is absent and the nasal crests are absent; the frontal bones on the skull are not separated by a midline anterior extension of the parietal bones; the anterior astragalar surface is flat; metacarpal I has a reduced distal medial condyle (noted by Ezcurra, 2006); the anterior margin of antorbital fossa is blunt and squared (noted by Carrano et al., 2012); the base of lacrimal vertical ramus width is less than 30% its height (noted by Carrano et al., 2012); the maxillary and dentary tooth rows end posteriorly at the anterior rim of the lacrimal bone (noted by Carrano et al., 2012)
Paleoecology
Provenance and occurrence
The holotype of C. rhodesiensis (QG1) has been recovered in Upper Elliot Formation in South Africa, as well as the Chitake River bonebed quarry at the Forest Sandstone Formation in Rhodesia (now known as Zimbabwe). In South Africa, several individuals were collected in 1985 from mudstone deposited during the Hettangian stage of the Jurassic period, approximately 201 to 199 million years ago.[8] In Zimbabwe, twenty-six individuals were collected in 1963, 1968 and 1972 from yellow sandstone deposited during the Hettangian stage of the Jurassic period, approximately 201 to 199 million years ago.[9][10][11]
Fauna and habitat
The Upper Elliot Formation is thought to have been an ancient floodplain. Fossils of the prosauropod dinosaur Massospondylus and Ignavusaurus have been recovered from the Upper Elliot Formation, which boasts the world's most diverse fauna of early Jurassic ornithischian dinosaurs, including Abrictosaurus, Fabrosaurus, Heterodontosaurus, and Lesothosaurus, among others. The Forest Sandstone Formation was the paleoenvironment of protosuchid crocodiles, sphenodonts, the dinosaur Massospondylus and indeterminate remains of a prosauropod. Paul (1988) noted that Megapnosaurus rhodesiensis lived among desert dunes and oases and hunted juvenile and adult prosauropods.[12]
Paleobiology
Appearance
C. rhodesiensis was a lean, elongated species of theropod dinosaur with an S shaped neck, long hind limbs that resembled the legs of large birds such as the Secretarybird, shorter forelimbs with four digits on each hand unlike most later theropods, and a long tail. While still lean, it sported a more robust frame than other Coelophysoidea members. Its extremely bird-like body lead to C. rhodesiensis being one of the first dinosaurs to be portrayed with feathers, though there is no direct evidence that it actually had feathers.[13]
Growth
Age determination studies using growth ring counts suggest that the longevity of C. rhodesiensis was approximately seven years.[14] Recent research has found that C. rhodesiensis had highly variable growth between individuals, with some specimens being larger in their immature phase than smaller adults were when completely mature; this indicates that the supposed presence of distinct morphs is simply the result of individual variation. This highly variable growth was likely ancestral to dinosaurs but later lost, and may have given such early dinosaurs an evolutionary advantage in surviving harsh environmental challenges.[15]
Feeding and diet
The supposed "weak joint" in the jaw, led to the early hypothesis that dinosaurs such as these were scavengers, as the front teeth and bone structure of the jaw were thought to be too weak to take down and hold struggling prey. C. rhodesiensis was one of the first dinosaurs to be portrayed with feathers, though there is no direct evidence that it actually had feathers. Paul (1988) noted that this genus, which may have hunted in packs, preyed upon prosauropods and early lizards.[12]
Comparisons between the scleral rings of C. rhodesiensis and modern birds and reptiles indicate that it may have been nocturnal.[16]
Paleopathology
In C. rhodesiensis, healed fractures of the tibia and metatarsus have been observed, but are very rare. "[T]he supporting butresses of the second sacral rib" in one specimen of Syntarsus rhodesiensis showed signs of fluctuating asymmetry. Fluctuating asymmetry results from developmental disturbances and is more common in populations under stress and can therefore be informative about the quality of conditions a dinosaur lived under.[17]
Ichnology
Dinosaur footprints that were later attributed to C. rhodesiensis were discovered in Rhodesia in 1915. These tracks were discovered at the Nyamandhlovu Sandstones Formation, in eolian red sandstone that was deposited in the Late Triassic, approximately 235 to 201 million years ago.[18]
References
- Rowe (1989). "A new species of the theropod dinosaur Syntarsus from the Early Jurassic Kayenta Formation of Arizona". Journal of Vertebrate Paleontology. 9 (2): 125–136. doi:10.1080/02724634.1989.10011748.
- Tykoski, 1998. The osteology of Syntarsus kayentakatae and its implications for ceratosaurid phylogeny. Unpublished Masters Thesis, University of Texas at Austin, 217 pp.
- Gay, 2010. Notes on Early Mesozoic theropods. Lulu Press. 44 pp.
- Mortimer, Mickey. "Coelophysoidea". Archived from the original on 4 May 2013. Retrieved 15 April 2013.
- Ezcurra, M. D.; Brusatte, S. L. (2011). "Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur Camposaurus arizonensis from the Late Triassic of North America". Palaeontology. 54 (4): 763–772. doi:10.1111/j.1475-4983.2011.01069.x.
- Raath, (1969). "A new Coelurosaurian dinosaur from the Forest Sandstone of Rhodesia." Arnoldia Rhodesia. 4 (28): 1-25.
- Tykoski, R. S. and Rowe, T., 2004, Ceratosauria, Chapter Three: In: The Dinosauria, Second Edition, edited by Weishampel, D.B., Dodson, P., and Osmolska, H., California University Press, p. 47-70.
- Munyikwa, D.; Raath, M. A. (1999). "Further material of the ceratosaurian dinosaur Syntarsus from the Elliot Formation (Early Jurassic) of South Africa". Palaeontologia Africana. 35: 55–59.
- M. A. Raath, 1969. A new coelurosaurian dinosaur from the Forest Sandstone of Rhodesia. Arnoldia (Rhodesia) 4(28):1-254
- Bond, G. (1965). "Some new fossil localities in the Karroo System of Rhodesia. Arnoldia, Series of Miscellaneous Publications". National Museum of Southern Rhodesia. 2 (11): 1–4.
- M. A. Raath, 1977. The Anatomy of the Triassic Theropod Syntarsus rhodesiensis (Saurischia: Podokesauridae) and a Consideration of Its Biology. Department of Zoology and Entomology, Rhodes University, Salisbury, Rhodesia 1-233
- Paul, G. S., 1988, Predatory Dinosaurs of the World, a complete Illustrated guide: New York Academy of sciences book, 464pp.
- Switek, Brian. (2013-04-16). My beloved Brontosaurus : on the road with old bones, new science, and our favorite dinosaurs (First ed.). New York. ISBN 9780374135065. OCLC 795174375.
- Chinsamy, A., (1994). Dinosaur bone histology: Implications and inferences. In Dino Fest (G. D. Rosenburg and D. L. Wolberg, Eds.), pp. 213-227. The Paleontological Society, Department of Geological Sciences, Univ. of Tennessee, Knoxville.
- Griffin, C.T.; Nesbitt, S.J. (2016). "Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds". Proceedings of the National Academy of Sciences of the United States of America. 113 (51): 14757–14762. doi:10.1073/pnas.1613813113. PMC 5187714. PMID 27930315.
- Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705–8. doi:10.1126/science.1200043. PMID 21493820.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
- Raath, M. A. (1972). "First record of dinosaur footprints from Rhodesia". Arnoldia. 5 (37): 1–5.