Haplogroup I-M438
Haplogroup I-M438, also known as I2 (and until 2007 as I1b), is a human DNA Y-chromosome haplogroup, a subclade of Haplogroup I-M170. Haplogroup I-M438 originated some time around 26,000–31,000 BCE. It originated in Europe and developed into several main subgroups : I2-M438*, I2a-L460, I2b-L415 and I2c-L596.[2] The haplogroup can be found all over Europe and reaches its maximum frequency in the Dinaric Alps (Balkans) via founder effect.[4] Examples of basal I-M438* have been found in males from Crete and Sicily .
| Haplogroup I-M438 | |
|---|---|
| Possible time of origin | 28–33,000 years ago[1] |
| Possible place of origin | South-Eastern or Eastern Europe. |
| Ancestor | I-M170 |
| Descendants | I-L460, I-L1251 |
| Defining mutations | M438/P215/S31 |
| Highest frequencies | I2a1a: Sardinia[2]
I2a1b: Bosnia and Herzegovina,[3] I2a2: Britain, Germany, and Sweden[2] |
Origin & prehistoric presence
Haplogroup I2a was the most frequent Y-DNA among western European mesolithic hunter gatherers (WHG) belonging to Villabruna Cluster. A 2015 study found haplogroup I2a in 13,500 year old remains from the Azilian culture (from Grotte du Bichon, modern Switzerland).[5] Subclades of I2a1 (I-P37.2), namely I-M423 and I-M26 have been found in remains of Western European Hunter-Gatherers dating from 10,000 to 8,000 years before present respectively.[6]
In a 2015 study published in Nature, the remains of six individuals from Motala ascribed to the Kongemose culture were successfully analyzed. With regards to Y-DNA, two individuals were ascribed to haplogroup I2a1b, one individual was ascribed to haplogroup I2a1, and one individual was ascribed to haplogroup I2c.[7]
Subclades
I-P37.2
The I-P37.2+, also known as I2a1 (or I-L68, is the SNP that defines I2a1. The subclade divergence for I-P37.2 occurred 10.7±4.8 kya. The age of YSTR variation for the P37.2 subclade is 8.0±4.0 kya.[2] It is the predominant version of I2 in Eastern Europe.[8] The I2a is further made up by sub-groups I-M26, I-M423, I-L1286, I-L880.
I-L158
Haplogroup I-M26 (or M26) has previously and is still sometimes called Eu-8, I1b2 (YCC), I1b1a, I2a1 or I2a2.
Haplogroup I-L158 (L158, L159.1/S169.1, M26) accounts for approximately 40% of all patrilines among the Sardinians.[9][10] It is also found at low to moderate frequency among populations of the Pyrenees (9.5% in Bortzerriak, Navarra; 9.7% in Chazetania, Aragon; 8% in Val d'Aran, Catalunya; 2.9% in Alt Urgell, Catalunya; and 8.1% in Baixa Cerdanya, Catalunya) and Iberia, and it has been found in 1.6% of a sample of Albanians living in the Republic of North Macedonia[11] and 1.2% (3/257) of a sample of Czechs.[12] The age of YSTR variation for the M26 subclade has been calculated at 8.0±4.0 kya.[2]
I-L178
I-L178 is very rare, but has been found in two persons from Germany and one from Poland. The age of YSTR variation for the M423 subclade is 8.8±3.6 kya.[1]
I-L621
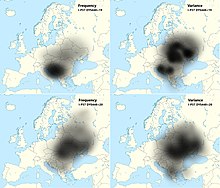
I2a1a2b-L621 is typical of the South Slavic populations of Southeastern Europe, being highest in Bosnia-Herzegovina (>50%) in Bosniaks, Croats, and Serbs,[3] because of which is often called as "Dinaric".[13] It has the highest variance and also high concentration in Eastern Europe (Ukraine, Southeastern Poland, Belarus).[14] According to YFull YTree it formed 11,400 YBP and had TMRCA 6,500 YBP, while its main subclades lineage is I-CTS10936 (6,500-5,600 YBP) > I-S19848 (5,600 YBP) > I-CTS4002 (5,600-5,100 YBP) > I-CTS10228 (5,100-3,400 YBP) > I-Y3120 (3,400-2,100 YBP) > I-Y18331 (2,100 YBP) / I-Z17855 (2,100-1650 YBP) / I-Y4460 (2,100 YBP) / I-S17250 (2,100-1,850 YBP) > I-PH908 (1,850-1,700 YBP).[15]
Older research considered that the high frequency of this subclade in the South Slavic-speaking populations to be the result of "pre-Slavic" paleolithic settlement in the region. Peričić et al. (2005) for instance place its expansion to have occurred "not earlier than the YD to Holocene transition and not later than the early Neolithic".[2][3][16][17] However the prehistoric autochthonous origin of the haplogroup I2 in the Balkans is now considered as out of date,[nb 1] as already Battaglia et al. (2009) observed highest variance of the haplogroup in Ukraine, and Zupan et al. (2013) noted that it suggests it arrived with Slavic migration from the homeland which was in present-day Ukraine.[22]
The research by O.M. Utevska (2017) concluded that the haplogroup STR haplotypes have the highest diversity in Ukraine, with ancestral STR marker result "DYS448=20" comprising "Dnieper-Carpathian" cluster, while younger derived result "DYS448=19" comprising the "Balkan cluster" which is predominant among the South Slavs.[14] This "Balkan cluster" also has the highest variance in Ukraine, which indicates that the very high frequency in the Western Balkan is because of a founder effect.[14] Utevska calculated that the STR cluster divergence and its secondary expansion from the middle reaches of the Dnieper river or from Eastern Carpathians towards the Balkan peninsula happened approximately 2,860 ± 730 years ago, relating it to the times before Slavs, but much after the decline of the Cucuteni–Trypillia culture.[14] More specifically, the "Balkan cluster" is represented by a single SNP, I-PH908, known as I2a1a2b1a1a1c in ISOGG phylogenetic tree (2019), and according to YFull YTree it formed and had TMRCA approximately 1,850-1,700 YBP (2nd-3rd century AD).[15]
It is considered that I-L621 could have been present in the Cucuteni–Trypillia culture,[23] but until now was only found G2a,[24] and another subclade I2a1a1-CTS595 was present in the Baden culture of the Calcholitic Carpathian Basin.[23][25] Although it is dominant among the modern Slavic peoples on the territory of the former Balkan provinces of the Roman Empire, until now it was not found among the samples from the Roman period and is almost absent in contemporary population of Italy.[13] It was found in the skeletal remains with artifacts, indicating leaders, of Hungarian conquerors of the Carpathian Basin from the 9th century, part of Western Eurasian-Slavic component of the Hungarians.[13][23] According to Fóthi et al. (2020), the distribution of ancestral subclades like of I-CTS10228 among contemporary carriers indicates a rapid expansion from Southeastern Poland, is mainly related to the Slavs, and the "largest demographic explosion occurred in the Balkans".[13] The earliest archeogenetic sample until now is Sungir 6 (~900 YBP) near Vladimir, Russia which belonged to the I-S17250 > I-Y5596 > I-Z16971 > I-Y5595 > I-A16681 subclade,[26][27] as well I-CTS10228 and I-Y3120 subclades were found in two Viking samples from Sweden (VK53) and Ukraine (VK542) with predominantly Slavic ancestry of which the second belongs to Gleb Svyatoslavich (11th century).[28]
I-M223
Haplogroup I2a2a (ISOGG 2014). The age of YSTR variation for the I-M223 subclade is 13.2±2.7 kya[2] and 12.3±3.1 kya.[1] YFull - "formed 15800 ybp, TMRCA 11700 ybp".[29] I-M223 has a peak in Germany and another in eastern Sweden, but also appears in Romania/Moldova, Russia, Greece, Italy and around the Black Sea.[30] Haplogroup I2a2a has been found in over 4% of the population only in Germany, the Netherlands, Belgium, Denmark, Scotland, and England (excluding Cornwall) – also the southern tips of Sweden and Norway in Northwest Europe; the provinces of Normandy, Maine, Anjou, and Perche in northwestern France; the province of Provence in southeastern France; the regions of Tuscany, Umbria, and Latium in Italy; Moldavia and the area around Russia's Ryazan Oblast and Mordovia in Eastern Europe. Of historical note, both haplogroups I-M253 and I-M223 appear at a low frequency in the historical regions of Bithynia and Galatia in Turkey. Haplogroup I2a2a also occurs among approximately 1% of Sardinians. The subclade divergence for M223 occurred 14.6±3.8 kya (Rootsi 2004).
Haplogroup I-M223 can be further subdivided in several subclades designated in the Y2012 ISOGG tree as follows:
- Haplogroup I-M223 with no further known polymorphisms,
- Haplogroup I-M284 defined by M284 polymorphism and including an undergroup Haplogroup I-L126 reserved for individuals derived for the L126/S165, L137/S166 polymorphisms, *Haplogroup I-L701 associated with L701 polymorphism, and Haplogroup I-Z161 denoting individuals derived for the Z161 polymorphism.
I-M284
Haplogroup I2a2a1a1 (ISOGG 2014). I-M284 has been found almost exclusively among the population of Great Britain, suggesting that the clade may have arisen in that island. I-M284 is comparatively rare in Ireland except in the north-east. In regard to north-east Ireland, the presence of this subclade "provides some tentative evidence of ancient flow with eastern areas that could support the idea that the La Tene culture was accompanied by some migration."[31] Where it is found in those of Irish descent with Gaelic surnames, this suggests an ancestor who arrived in Ireland from Celtic Britain.[31] Men with several Gaelic surnames such as McGuinness and McCartan bear this subclade, family groups that have a historically recorded 6th-century common ancestor, thus it is not the result of known recent gene flow between Britain and Ireland.[31] While subclades of I-M284 are atypical of Ireland they are relatively common in continental Europe.[31] The observed mutational divergence between men with this subclade suggests its foundation very approximately at 300 BC, thus dates and geography are circumstantially but not securely associated with Iron Age continental Europe.[31]
I-CTS10057
Continentals. Mother Haplogroup for group I-Z161 (Continental 1 and 2) and I-L701 group (Continental 3). Around 10.000 years old.
I-Z161
Haplogroup I2a2a1b2 (ISOGG 2014). Z161+ defines the I2 Continental clade (except Continental 3). Its age is estimated around 7,000 years old. It is mainly found in North Europe, especially in Denmark, Germany, the Netherlands, and England. In Northwest Sicily it can also be found; this is believed to be due to remnants of a Norman settlement.
I-L701
Called Continental 3. Continental 3 has a wide distribution. Found in Central Europe from Germany, Austria to Poland, Romania and Ukraine, but also in lower frequencies in Greece, Italy, France, Spain, England, Ireland, and Armenia. It may have been disseminated in part by the Goths. It is nearly absent from Scandinavia and Scotland.
Subclades list
| Haplogroup I |
| ||||||||||||||||||||||||||||||
Subclades list |
|---|
|
|
See also
- Human Y-chromosome DNA haplogroups
- Haplogroup I (Y-DNA)
- Haplogroup I1 (Y-DNA)
References
- Underhill, Peter A.; Myres, Natalie M.; Rootsi, Siiri; Chow, Cheryl-Emiliane T.; Lin, Alice A.; Otillar, Robert P.; King, Roy; Zhivotovsky, Lev A.; Balanovsky, Oleg; Pshenichnov, Andrey; Ritchie, Kathleen H.; Cavalli-Sforza, L. Luca; Kivisild, Toomas; Villems, Richard; Woodward, Scott R. (2007). "New phylogenetic relationships for Y-chromosome haplogroup I: Reappraising its Phylogeography and Prehistory". In Mellars, Paul; Boyle, Katie; Bar-Yosef, Ofer; Stringer, Chris (eds.). Rethinking the Human Evolution. pp. 33–42. ISBN 978-1-902937-46-5.
- Rootsi, Siiri; Magri, Chiara; Kivisild, Toomas; Benuzzi, Giorgia; Help, Hela; Bermisheva, Marina; Kutuev, Ildus; Barać, Lovorka; Peričić, Marijana; Balanovsky, Oleg; Pshenichnov, Andrey; Dion, Daniel; Grobei, Monica; Zhivotovsky, Lev A.; Battaglia, Vincenza; Achilli, Alessandro; Al-Zahery, Nadia; Parik, Jüri; King, Roy; Cinnioğlu, Cengiz; Khusnutdinova, Elsa; Rudan, Pavao; Balanovska, Elena; Scheffrahn, Wolfgang; Simonescu, Maya; Brehm, Antonio; Goncalves, Rita; Rosa, Alexandra; Moisan, Jean-Paul; Chaventre, Andre; Ferak, Vladimir; Füredi, Sandor; Oefner, Peter J.; Shen, Peidong; Beckman, Lars; Mikerezi, Ilia; Terzić, Rifet; Primorac, Dragan; Cambon-Thomsen, Anne; Krumina, Astrida; Torroni, Antonio; Underhill, Peter A.; Santachiara-Benerecetti, A. Silvana; Villems, Richard; Semino, Ornella (2004). "Phylogeography of Y-Chromosome Haplogroup I Reveals Distinct Domains of Prehistoric Gene Flow in Europe". American Journal of Human Genetics. 75 (1): 128–137. doi:10.1086/422196. PMC 1181996. PMID 15162323.
- Peričić, Marijana; Lauc, Lovorka Barać; Klarić, Irena Martinović; Rootsi, Siiri; Janićijević, Branka; Rudan, Igor; Terzić, Rifet; Čolak, Ivanka; Kvesić, Ante; Popović, Dan; Šijački, Ana; Behluli, Ibrahim; Đorđević, Dobrivoje; Efremovska, Ljudmila; Bajec, Đorđe D.; Stefanović, Branislav D.; Villems, Richard; Rudan, Pavao (October 2005). "High-Resolution Phylogenetic Analysis of Southeastern Europe Traces Major Episodes of Paternal Gene Flow Among Slavic Populations". Molecular Biology and Evolution. 22 (10): 1964–1975. doi:10.1093/molbev/msi185. PMID 15944443.
- Grasgruber, Pavel; Popović, Stevo; Bokuvka, Dominik; Davidović, Ivan; Hřebíčková, Sylva; Ingrová, Pavlína; Potpara, Predrag; Prce, Stipan; Stračárová, Nikola (12 April 2017). "The mountains of giants: an anthropometric survey of male youths in Bosnia and Herzegovina". Royal Society Open Science. 4 (4): 161054. Bibcode:2017RSOS....461054G. doi:10.1098/rsos.161054. PMC 5414258. PMID 28484621.
- Jones, Eppie R.; Gonzalez-Fortes, Gloria; Connell, Sarah; Siska, Veronika; Eriksson, Anders; Martiniano, Rui; McLaughlin, Russell L.; Gallego Llorente, Marcos; Cassidy, Lara M.; Gamba, Cristina; Meshveliani, Tengiz; Bar-Yosef, Ofer; Müller, Werner; Belfer-Cohen, Anna; Matskevich, Zinovi; Jakeli, Nino; Higham, Thomas F. G.; Currat, Mathias; Lordkipanidze, David; Hofreiter, Michael; Manica, Andrea; Pinhasi, Ron; Bradley, Daniel G. (2015). "Upper Palaeolithic genomes reveal deep roots of modern Eurasians". Nature Communications. 6: 8912. Bibcode:2015NatCo...6.8912J. doi:10.1038/ncomms9912. PMC 4660371. PMID 26567969.
- Archived 2017-04-30 at the Wayback Machine Mesolithic Western Eurasian DNA
- Mathieson, Iain (December 24, 2015). "Genome-wide patterns of selection in 230 ancient Eurasians". Nature. 528 (7583): 499–503. Bibcode:2015Natur.528..499M. doi:10.1038/nature16152. PMC 4918750. PMID 26595274.CS1 maint: ref=harv (link)
- Sazzini, Marco; Sarno, Stefania; Luiselli, Donata (2014). "The Mediterranean Human Population: An Anthropological Genetics Perspective". The Mediterranean Sea. pp. 529–551. doi:10.1007/978-94-007-6704-1_31. ISBN 978-94-007-6703-4.
- Rootsi, Siiri (31 December 2006). "Y-Chromosome haplogroup I prehistoric gene flow in Europe". Documenta Praehistorica. 33: 17–20. doi:10.4312/dp.33.3.
- Francalacci, Paolo; Morelli, Laura; Angius, Andrea; Berutti, Riccardo; Reinier, Frederic; Atzeni, Rossano; Pilu, Rosella; Busonero, Fabio; Maschio, Andrea; Zara, Ilenia; Sanna, Daria; Useli, Antonella; Urru, Maria Francesca; Marcelli, Marco; Cusano, Roberto; Oppo, Manuela; Zoledziewska, Magdalena; Pitzalis, Maristella; Deidda, Francesca; Porcu, Eleonora; Poddie, Fausto; Kang, Hyun Min; Lyons, Robert; Tarrier, Brendan; Gresham, Jennifer Bragg; Li, Bingshan; Tofanelli, Sergio; Alonso, Santos; Dei, Mariano; Lai, Sandra; Mulas, Antonella; Whalen, Michael B.; Uzzau, Sergio; Jones, Chris; Schlessinger, David; Abecasis, Gonçalo R.; Sanna, Serena; Sidore, Carlo; Cucca, Francesco (2 August 2013). "Low-Pass DNA Sequencing of 1200 Sardinians Reconstructs European Y-Chromosome Phylogeny". Science. 341 (6145): 565–569. Bibcode:2013Sci...341..565F. doi:10.1126/science.1237947. PMC 5500864. PMID 23908240.
- Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri; Marjanovic, Damir; Primorac, Dragan; Hadziselimovic, Rifat; Vidovic, Stojko; Drobnic, Katia; Durmishi, Naser; Torroni, Antonio; Santachiara-Benerecetti, A Silvana; Underhill, Peter A; Semino, Ornella (June 2009). "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe". European Journal of Human Genetics. 17 (6): 820–830. doi:10.1038/ejhg.2008.249. PMC 2947100. PMID 19107149.
- Luca, F.; Di Giacomo, F.; Benincasa, T.; Popa, L.O.; Banyko, J.; Kracmarova, A.; Malaspina, P.; Novelletto, A.; Brdicka, R. (January 2007). "Y-chromosomal variation in the Czech Republic". American Journal of Physical Anthropology. 132 (1): 132–139. doi:10.1002/ajpa.20500. hdl:2108/35058. PMID 17078035.
- Fóthi, E.; Gonzalez, A.; Fehér, T.; et al. (2020), "Genetic analysis of male Hungarian Conquerors: European and Asian paternal lineages of the conquering Hungarian tribes", Archaeological and Anthropological Sciences, 12 (1), doi:10.1007/s12520-019-00996-0
- O.M. Utevska (2017). Генофонд українців за різними системами генетичних маркерів: походження і місце на європейському генетичному просторі [The gene pool of Ukrainians revealed by different systems of genetic markers: the origin and statement in Europe] (PhD) (in Ukrainian). National Research Center for Radiation Medicine of National Academy of Sciences of Ukraine. pp. 219–226, 302.
- "I-PH908 YTree v8.06.01". YFull.com. 27 June 2020. Retrieved 17 July 2020.
- Marjanovic, D.; Fornarino, S.; Montagna, S.; Primorac, D.; Hadziselimovic, R.; Vidovic, S.; Pojskic, N.; Battaglia, V.; Achilli, A.; Drobnic, K.; Andjelinovic, S.; Torroni, A.; Santachiara-Benerecetti, A. S.; Semino, O. (November 2005). "The Peopling of Modern Bosnia-Herzegovina: Y-chromosome Haplogroups in the Three Main Ethnic Groups". Annals of Human Genetics. 69 (6): 757–763. doi:10.1111/j.1529-8817.2005.00190.x. PMID 16266413.
- Rębała, Krzysztof; Mikulich, Alexei I.; Tsybovsky, Iosif S.; Siváková, Daniela; Džupinková, Zuzana; Szczerkowska-Dobosz, Aneta; Szczerkowska, Zofia (16 March 2007). "Y-STR variation among Slavs: evidence for the Slavic homeland in the middle Dnieper basin". Journal of Human Genetics. 52 (5): 406–414. doi:10.1007/s10038-007-0125-6. PMID 17364156.
- "I-P37 YTree v8.06.01". YFull.com. 27 June 2020. Retrieved 17 July 2020.
- "I2a Y-Haplogroup - Results: I2a2a-Dinaric". Family Tree DNA. Retrieved 11 November 2018.
Ken Nordtvedt has split I2a2-M423-Dinaric into Din-N and Din-S. Din-N is older than Din-S. N=north of the Danube and S=south of the Danube River ... May 8, 2007: Dinaric I1b1 and DYS 448. DYS448 19 for S and 20 for N.
- Bernie Cullen (22 August 2016). "Link to I-L621 tree showing major STR clusters (Updated)". i2aproject.blogspot.com. Blogger. Retrieved 3 April 2019.
- "Y-DNA Haplogroup I and its Subclades - 2019-2020". ISOGG. 1 October 2019. Retrieved 17 July 2020.
- A. Zupan; et al. (2013). "The paternal perspective of the Slovenian population and its relationship with other populations". Annals of Human Biology. 40 (6): 515–526. doi:10.3109/03014460.2013.813584. PMID 23879710.
- Neparáczki, Endre; et al. (2019). "Y-chromosome haplogroups from Hun, Avar and conquering Hungarian period nomadic people of the Carpathian Basin". Scientific Reports. Nature Research. 9 (16569): 16569. doi:10.1038/s41598-019-53105-5. PMC 6851379. PMID 31719606.
- Mathieson, Iain (February 21, 2018). "The Genomic History of Southeastern Europe". Nature. 555 (7695): 197–203. doi:10.1038/nature25778. PMC 6091220. PMID 29466330.
- Lipson, Mark (2017). "Parallel ancient genomic transects reveal complex population history of early European farmers". Nature. 551: 368–372. doi:10.1038/nature24476.
- Sikora, Martin; Seguin-Orlando, Andaine; Sousa, Vitor C; Albrechtsen, Anders; Korneliussen, Thorfinn; Ko, Amy; Rasmussen, Simon; Dupanloup, Isabelle; Nigst, Philip R; Bosch, Marjolein D; Renaud, Gabriel; Allentoft, Morten E; Margaryan, Ashot; Vasilyev, Sergey V; Veselovskaya, Elizaveta V; Borutskaya, Svetlana B; Deviese, Thibaut; Comeskey, Dan; Higham, Tom; Manica, Andrea; Foley, Robert; Meltzer, David J; Nielsen, Rasmus; Excoffier, Laurent; Mirazon Lahr, Marta; Orlando, Ludovic; Willerslev, Eske (2017). "Ancient genomes show social and reproductive behavior of early Upper Paleolithic foragers". Science. 358 (6363): 659–662. doi:10.1126/science.aao1807. PMID 28982795.
- "I-A16681 YTree v8.06.01". YFull.com. 27 June 2020. Retrieved 17 July 2020.
- Margaryan, Ashot (17 July 2019). "Population genomics of the Viking world". bioRxiv 10.1101/703405. Cite journal requires
|journal=(help) - "I-M223 YTree".
- Chiaroni, J.; Underhill, P. A.; Cavalli-Sforza, L. L. (17 November 2009). "Y chromosome diversity, human expansion, drift, and cultural evolution". Proceedings of the National Academy of Sciences. 106 (48): 20174–20179. Bibcode:2009PNAS..10620174C. doi:10.1073/pnas.0910803106. PMC 2787129. PMID 19920170.
- McEvoy, Brian P.; Bradly, Daniel G. (2010). "Irish Genetics and Celts". In Cunliffe, Barry W.; Koch, John T. (eds.). Celtic from the West: Alternative Perspectives from Archaeology, Genetics, Language, and Literature. Oxbow Books. pp. 107–120. ISBN 978-1-84217-410-4.
- The SNP I-P37 itself formed approximately 21,000 YBP and had TMRCA 18,400 YBP according to YFull YTree,[18] being too old and widespread as an SNP for argumentation of ancient autochthony or medieval migration as well the old research used outdated nomenclature. According to "I-P37 (I2a)" project at Family Tree DNA, the divergence at STR marker DYS448 20 > 19 is reported since 2007,[19] while the SNP which defines the STR Dinaric-South cluster, I-PH908, is reported since 2014.[20] The SNP I-PH908 at ISOGG phylogenetic tree is named as I2a1a2b1a1a1c,[21] while formed and had TMRCA approximately 1,800 YBP according to YFull.[15]
External links
Relationship to haplogroups and subclades
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||