Y chromosome
The Y chromosome is one of two sex chromosomes (allosomes) in mammals, including humans, and many other animals. The other is the X chromosome. Y is normally the sex-determining chromosome in many species, since it is the presence or absence of Y that typically determines the male or female sex of offspring produced in sexual reproduction. In mammals, the Y chromosome contains the gene SRY, which triggers male development. The DNA in the human Y chromosome is composed of about 59 million base pairs.[5] The Y chromosome is passed only from father to son. With a 30% difference between humans and chimpanzees, the Y chromosome is one of the fastest-evolving parts of the human genome.[6] The human Y chromosome carries an estimated 100-200 genes, with between 45 and 73 of these protein-coding. All single-copy Y-linked genes are hemizygous (present on only one chromosome) except in cases of aneuploidy such as XYY syndrome or XXYY syndrome.
| Human Y chromosome | |
|---|---|
Human Y chromosome (after G-banding) | |
 Y chromosome in human male karyogram | |
| Features | |
| Length (bp) | 57,227,415 bp (GRCh38)[1] |
| No. of genes | 63 (CCDS)[2] |
| Type | Allosome |
| Centromere position | Acrocentric[3] (10.4 Mbp[4]) |
| Complete gene lists | |
| CCDS | Gene list |
| HGNC | Gene list |
| UniProt | Gene list |
| NCBI | Gene list |
| External map viewers | |
| Ensembl | Chromosome Y |
| Entrez | Chromosome Y |
| NCBI | Chromosome Y |
| UCSC | Chromosome Y |
| Full DNA sequences | |
| RefSeq | NC_000024 (FASTA) |
| GenBank | CM000686 (FASTA) |
Overview
Discovery
The Y chromosome was identified as a sex-determining chromosome by Nettie Stevens at Bryn Mawr College in 1905 during a study of the mealworm Tenebrio molitor. Edmund Beecher Wilson independently discovered the same mechanisms the same year. Stevens proposed that chromosomes always existed in pairs and that the Y chromosome was the pair of the X chromosome discovered in 1890 by Hermann Henking. She realized that the previous idea of Clarence Erwin McClung, that the X chromosome determines sex, was wrong and that sex determination is, in fact, due to the presence or absence of the Y chromosome. Stevens named the chromosome "Y" simply to follow on from Henking's "X" alphabetically.[7][8]
The idea that the Y chromosome was named after its similarity in appearance to the letter "Y" is mistaken. All chromosomes normally appear as an amorphous blob under the microscope and only take on a well-defined shape during mitosis. This shape is vaguely X-shaped for all chromosomes. It is entirely coincidental that the Y chromosome, during mitosis, has two very short branches which can look merged under the microscope and appear as the descender of a Y-shape.[9]
Variations
Most therian mammals have only one pair of sex chromosomes in each cell. Males have one Y chromosome and one X chromosome, while females have two X chromosomes. In mammals, the Y chromosome contains a gene, SRY, which triggers embryonic development as a male. The Y chromosomes of humans and other mammals also contain other genes needed for normal sperm production.
There are exceptions, however. Among humans, some men have two Xs and a Y ("XXY", see Klinefelter syndrome), or one X and two Ys (see XYY syndrome), and some women have three Xs or a single X instead of a double X ("X0", see Turner syndrome). There are other exceptions in which SRY is damaged (leading to an XY female), or copied to the X (leading to an XX male).
Origins and evolution
Before Y chromosome
Many ectothermic vertebrates have no sex chromosomes. If they have different sexes, sex is determined environmentally rather than genetically. For some of them, especially reptiles, sex depends on the incubation temperature. Some vertebrates are hermaphrodites, although other than a very few ray-finned fish, they are sequential (the same organism produces male or female gametes, but never both, at different points in its life), rather than simultaneous (the same organism producing both male and female gametes at the same time).
Origin
The X and Y chromosomes are thought to have evolved from a pair of identical chromosomes,[10][11] termed autosomes, when an ancestral animal developed an allelic variation, a so-called "sex locus" – simply possessing this allele caused the organism to be male.[12] The chromosome with this allele became the Y chromosome, while the other member of the pair became the X chromosome. Over time, genes that were beneficial for males and harmful to (or had no effect on) females either developed on the Y chromosome or were acquired through the process of translocation.[13]
Until recently, the X and Y chromosomes were thought to have diverged around 300 million years ago.[14] However, research published in 2010,[15] and particularly research published in 2008 documenting the sequencing of the platypus genome,[16] has suggested that the XY sex-determination system would not have been present more than 166 million years ago, at the split of the monotremes from other mammals.[17] This re-estimation of the age of the therian XY system is based on the finding that sequences that are on the X chromosomes of marsupials and eutherian mammals are present on the autosomes of platypus and birds.[17] The older estimate was based on erroneous reports that the platypus X chromosomes contained these sequences.[18][19]
Recombination inhibition
Recombination between the X and Y chromosomes proved harmful—it resulted in males without necessary genes formerly found on the Y chromosome, and females with unnecessary or even harmful genes previously only found on the Y chromosome. As a result, genes beneficial to males accumulated near the sex-determining genes, and recombination in this region was suppressed in order to preserve this male specific region.[12] Over time, the Y chromosome changed in such a way as to inhibit the areas around the sex determining genes from recombining at all with the X chromosome. As a result of this process, 95% of the human Y chromosome is unable to recombine. Only the tips of the Y and X chromosomes recombine. The tips of the Y chromosome that could recombine with the X chromosome are referred to as the pseudoautosomal region. The rest of the Y chromosome is passed on to the next generation intact, allowing for its use in tracking human evolution.
Degeneration
By one estimate, the human Y chromosome has lost 1,393 of its 1,438 original genes over the course of its existence, and linear extrapolation of this 1,393-gene loss over 300 million years gives a rate of genetic loss of 4.6 genes per million years.[20] Continued loss of genes at the rate of 4.6 genes per million years would result in a Y chromosome with no functional genes – that is the Y chromosome would lose complete function – within the next 10 million years, or half that time with the current age estimate of 160 million years.[12][21] Comparative genomic analysis reveals that many mammalian species are experiencing a similar loss of function in their heterozygous sex chromosome. Degeneration may simply be the fate of all non-recombining sex chromosomes, due to three common evolutionary forces: high mutation rate, inefficient selection, and genetic drift.[12]
However, comparisons of the human and chimpanzee Y chromosomes (first published in 2005) show that the human Y chromosome has not lost any genes since the divergence of humans and chimpanzees between 6–7 million years ago,[22] and a scientific report in 2012 stated that only one gene had been lost since humans diverged from the rhesus macaque 25 million years ago.[23] These facts provide direct evidence that the linear extrapolation model is flawed and suggest that the current human Y chromosome is either no longer shrinking or is shrinking at a much slower rate than the 4.6 genes per million years estimated by the linear extrapolation model.
High mutation rate
The human Y chromosome is particularly exposed to high mutation rates due to the environment in which it is housed. The Y chromosome is passed exclusively through sperm, which undergo multiple cell divisions during gametogenesis. Each cellular division provides further opportunity to accumulate base pair mutations. Additionally, sperm are stored in the highly oxidative environment of the testis, which encourages further mutation. These two conditions combined put the Y chromosome at a greater opportunity of mutation than the rest of the genome.[12] The increased mutation opportunity for the Y chromosome is reported by Graves as a factor 4.8.[12] However, her original reference obtains this number for the relative mutation rates in male and female germ lines for the lineage leading to humans.[24]
The observation that the Y chromosome experiences little meiotic recombination and has an accelerated rate of mutation and degradative change compared to the rest of the genome suggests an evolutionary explanation for the adaptive function of meiosis with respect to the main body of genetic information. Brandeis[25] proposed that the basic function of meiosis (particularly meiotic recombination) is the conservation of the integrity of the genome, a proposal consistent with the idea that meiosis is an adaptation for repairing DNA damage.[26]
Inefficient selection
Without the ability to recombine during meiosis, the Y chromosome is unable to expose individual alleles to natural selection. Deleterious alleles are allowed to "hitchhike" with beneficial neighbors, thus propagating maladapted alleles into the next generation. Conversely, advantageous alleles may be selected against if they are surrounded by harmful alleles (background selection). Due to this inability to sort through its gene content, the Y chromosome is particularly prone to the accumulation of "junk" DNA. Massive accumulations of retrotransposable elements are scattered throughout the Y.[12] The random insertion of DNA segments often disrupts encoded gene sequences and renders them nonfunctional. However, the Y chromosome has no way of weeding out these "jumping genes". Without the ability to isolate alleles, selection cannot effectively act upon them.
A clear, quantitative indication of this inefficiency is the entropy rate of the Y chromosome. Whereas all other chromosomes in the human genome have entropy rates of 1.5–1.9 bits per nucleotide (compared to the theoretical maximum of exactly 2 for no redundancy), the Y chromosome's entropy rate is only 0.84.[27] This means the Y chromosome has a much lower information content relative to its overall length; it is more redundant.
Genetic drift
Even if a well adapted Y chromosome manages to maintain genetic activity by avoiding mutation accumulation, there is no guarantee it will be passed down to the next generation. The population size of the Y chromosome is inherently limited to 1/4 that of autosomes: diploid organisms contain two copies of autosomal chromosomes while only half the population contains 1 Y chromosome. Thus, genetic drift is an exceptionally strong force acting upon the Y chromosome. Through sheer random assortment, an adult male may never pass on his Y chromosome if he only has female offspring. Thus, although a male may have a well adapted Y chromosome free of excessive mutation, it may never make it into the next gene pool.[12] The repeat random loss of well-adapted Y chromosomes, coupled with the tendency of the Y chromosome to evolve to have more deleterious mutations rather than less for reasons described above, contributes to the species-wide degeneration of Y chromosomes through Muller's ratchet.[28]
Gene conversion
As it has been already mentioned, the Y chromosome is unable to recombine during meiosis like the other human chromosomes; however, in 2003, researchers from MIT discovered a process which may slow down the process of degradation. They found that human Y chromosome is able to "recombine" with itself, using palindrome base pair sequences.[29] Such a "recombination" is called gene conversion.
In the case of the Y chromosomes, the palindromes are not noncoding DNA; these strings of bases contain functioning genes important for male fertility. Most of the sequence pairs are greater than 99.97% identical. The extensive use of gene conversion may play a role in the ability of the Y chromosome to edit out genetic mistakes and maintain the integrity of the relatively few genes it carries. In other words, since the Y chromosome is single, it has duplicates of its genes on itself instead of having a second, homologous, chromosome. When errors occur, it can use other parts of itself as a template to correct them.
Findings were confirmed by comparing similar regions of the Y chromosome in humans to the Y chromosomes of chimpanzees, bonobos and gorillas. The comparison demonstrated that the same phenomenon of gene conversion appeared to be at work more than 5 million years ago, when humans and the non-human primates diverged from each other.
Future evolution
In the terminal stages of the degeneration of the Y chromosome, other chromosomes increasingly take over genes and functions formerly associated with it. Finally, the Y chromosome disappears entirely, and a new sex-determining system arises.[12] Several species of rodent in the sister families Muridae and Cricetidae have reached these stages,[30][31] in the following ways:
- The Transcaucasian mole vole, Ellobius lutescens, the Zaisan mole vole, Ellobius tancrei, and the Japanese spinous country rats Tokudaia osimensis and Tokudaia tokunoshimensis, have lost the Y chromosome and SRY entirely.[12][32][33] Tokudaia spp. have relocated some other genes ancestrally present on the Y chromosome to the X chromosome.[33] Both sexes of Tokudaia spp. and Ellobius lutescens have an XO genotype (Turner syndrome),[33] whereas all Ellobius tancrei possess an XX genotype.[12] The new sex-determining system(s) for these rodents remains unclear.
- The wood lemming Myopus schisticolor, the Arctic lemming, Dicrostonyx torquatus, and multiple species in the grass mouse genus Akodon have evolved fertile females who possess the genotype generally coding for males, XY, in addition to the ancestral XX female, through a variety of modifications to the X and Y chromosomes.[30][34][35]
- In the creeping vole, Microtus oregoni, the females, with just one X chromosome each, produce X gametes only, and the males, XY, produce Y gametes, or gametes devoid of any sex chromosome, through nondisjunction.[36]
Outside of the rodents, the black muntjac, Muntiacus crinifrons, evolved new X and Y chromosomes through fusions of the ancestral sex chromosomes and autosomes.[37]
1:1 sex ratio
Fisher's principle outlines why almost all species using sexual reproduction have a sex ratio of 1:1. W. D. Hamilton gave the following basic explanation in his 1967 paper on "Extraordinary sex ratios",[38] given the condition that males and females cost equal amounts to produce:
- Suppose male births are less common than female.
- A newborn male then has better mating prospects than a newborn female, and therefore can expect to have more offspring.
- Therefore, parents genetically disposed to produce males tend to have more than average numbers of grandchildren born to them.
- Therefore, the genes for male-producing tendencies spread, and male births become more common.
- As the 1:1 sex ratio is approached, the advantage associated with producing males dies away.
- The same reasoning holds if females are substituted for males throughout. Therefore, 1:1 is the equilibrium ratio.
Non-therian Y chromosome
Many groups of organisms in addition to therian mammals have Y chromosomes, but these Y chromosomes do not share common ancestry with therian Y chromosomes. Such groups include monotremes, Drosophila, some other insects, some fish, some reptiles, and some plants. In Drosophila melanogaster, the Y chromosome does not trigger male development. Instead, sex is determined by the number of X chromosomes. The D. melanogaster Y chromosome does contain genes necessary for male fertility. So XXY D. melanogaster are female, and D. melanogaster with a single X (X0), are male but sterile. There are some species of Drosophila in which X0 males are both viable and fertile.
ZW chromosomes
Other organisms have mirror image sex chromosomes: where the homogeneous sex is the male, said to have two Z chromosomes, and the female is the heterogeneous sex, and said to have a Z chromosome and a W chromosome. For example, female birds, snakes, and butterflies have ZW sex chromosomes, and males have ZZ sex chromosomes.
Non-inverted Y chromosome
There are some species, such as the Japanese rice fish, the XY system is still developing and cross over between the X and Y is still possible. Because the male specific region is very small and contains no essential genes, it is even possible to artificially induce XX males and YY females to no ill effect.[39]
Multiple XY pairs
Monotremes possess four or five (platypus) pairs of XY sex chromosomes, each pair consisting of sex chromosomes with homologous regions. The chromosomes of neighboring pairs are partially homologous, such that a chain is formed during mitosis.[18] The first X chromosome in the chain is also partially homologous with the last Y chromosome, indicating that profound rearrangements, some adding new pieces from autosomes, have occurred in history.[40][41](fig. 5)
Platypus sex chromosomes have strong sequence similarity with the avian Z chromosome, (indicating close homology),[16] and the SRY gene so central to sex-determination in most other mammals is apparently not involved in platypus sex-determination.[17]
Human Y chromosome
In humans, the Y chromosome spans about 58 million base pairs (the building blocks of DNA) and represents almost 2% of the total DNA in a male cell.[42] The human Y chromosome contains over 200 genes, at least 72 of which code for proteins.[5] Traits that are inherited via the Y chromosome are called Y-linked traits, or holandric traits (from Ancient Greek ὅλος hólos, "whole" + ἀνδρός andrós, "male").[43]
Men can lose the Y chromosome in a subset of cells, which is called the mosaic loss of chromosome Y (LOY). This post-zygotic mutation is strongly associated with age, affecting about 15% of men 70 years of age. Smoking is another important risk factor for LOY.[44] It has been found that men with a higher percentage of hematopoietic stem cells in blood lacking the Y chromosome (and perhaps a higher percentage of other cells lacking it) have a higher risk of certain cancers and have a shorter life expectancy. Men with LOY (which was defined as no Y in at least 18% of their hematopoietic cells) have been found to die 5.5 years earlier on average than others. This has been interpreted as a sign that the Y chromosome plays a role going beyond sex determination and reproduction[45] (although the loss of Y may be an effect rather than a cause). Male smokers have between 1.5 and 2 times the risk of non-respiratory cancers as female smokers.[46][47]
Non-combining region of Y (NRY)
The human Y chromosome is normally unable to recombine with the X chromosome, except for small pieces of pseudoautosomal regions at the telomeres (which comprise about 5% of the chromosome's length). These regions are relics of ancient homology between the X and Y chromosomes. The bulk of the Y chromosome, which does not recombine, is called the "NRY", or non-recombining region of the Y chromosome.[48] The single-nucleotide polymorphisms (SNPs) in this region are used to trace direct paternal ancestral lines.
Genes
Number of genes
The following are some of the gene count estimates of human Y chromosome. Because researchers use different approaches to genome annotation their predictions of the number of genes on each chromosome varies (for technical details, see gene prediction). Among various projects, the collaborative consensus coding sequence project (CCDS) takes an extremely conservative strategy. So CCDS's gene number prediction represents a lower bound on the total number of human protein-coding genes.[49]
| Estimated by | Protein-coding genes | Non-coding RNA genes | Pseudogenes | Source | Release date |
|---|---|---|---|---|---|
| CCDS | 63 | — | — | [2] | 2016-09-08 |
| HGNC | 45 | 55 | 381 | [50] | 2017-05-12 |
| Ensembl | 63 | 109 | 392 | [51] | 2017-03-29 |
| UniProt | 47 | — | — | [52] | 2018-02-28 |
| NCBI | 73 | 122 | 400 | [53][54][55] | 2017-05-19 |
Gene list
In general, the human Y chromosome is extremely gene poor—it is one of the largest gene deserts in the human genome. Disregarding pseudoautosomal genes, genes encoded on the human Y chromosome include:
- NRY, with corresponding gene on X chromosome
- NRY, other
- AZF1 (azoospermia factor 1)
- BPY2 (basic protein on the Y chromosome)
- DAZ1 (deleted in azoospermia)
- DAZ2
- DFNY1 encoding protein Deafness, Y-linked 1
- PRKY (protein kinase, Y-linked)
- RBMY1A1
- SRY (sex-determining region)
- TSPY (testis-specific protein)
- USP9Y
- UTY (ubiquitously transcribed TPR gene on Y chromosome)
- ZFY (zinc finger protein)
Y-chromosome-linked diseases
Diseases linked to the Y chromosome typically involve an aneuploidy, an atypical number of chromosomes.
Y chromosome microdeletion
Y chromosome microdeletion (YCM) is a family of genetic disorders caused by missing genes in the Y chromosome. Many affected men exhibit no symptoms and lead normal lives. However, YCM is also known to be present in a significant number of men with reduced fertility or reduced sperm count.
Defective Y chromosome
This results in the person presenting a female phenotype (i.e., is born with female-like genitalia) even though that person possesses an XY karyotype. The lack of the second X results in infertility. In other words, viewed from the opposite direction, the person goes through defeminization but fails to complete masculinization.
The cause can be seen as an incomplete Y chromosome: the usual karyotype in these cases is 45X, plus a fragment of Y. This usually results in defective testicular development, such that the infant may or may not have fully formed male genitalia internally or externally. The full range of ambiguity of structure may occur, especially if mosaicism is present. When the Y fragment is minimal and nonfunctional, the child is usually a girl with the features of Turner syndrome or mixed gonadal dysgenesis.
XXY
Klinefelter syndrome (47, XXY) is not an aneuploidy of the Y chromosome, but a condition of having an extra X chromosome, which usually results in defective postnatal testicular function. The mechanism is not fully understood; it does not seem to be due to direct interference by the extra X with expression of Y genes.
XYY
47, XYY syndrome (simply known as XYY syndrome) is caused by the presence of a single extra copy of the Y chromosome in each of a male's cells. 47, XYY males have one X chromosome and two Y chromosomes, for a total of 47 chromosomes per cell. Researchers have found that an extra copy of the Y chromosome is associated with increased stature and an increased incidence of learning problems in some boys and men, but the effects are variable, often minimal, and the vast majority do not know their karyotype.[58]
In 1965 and 1966 Patricia Jacobs and colleagues published a chromosome survey of 315 male patients at Scotland's only special security hospital for the developmentally disabled, finding a higher than expected number of patients to have an extra Y chromosome.[59] The authors of this study wondered "whether an extra Y chromosome predisposes its carriers to unusually aggressive behaviour", and this conjecture "framed the next fifteen years of research on the human Y chromosome".[60]
Through studies over the next decade, this conjecture was shown to be incorrect: the elevated crime rate of XYY males is due to lower median intelligence and not increased aggression,[61] and increased height was the only characteristic that could be reliably associated with XYY males.[62] The "criminal karyotype" concept is therefore inaccurate.[58]
Rare
The following Y-chromosome-linked diseases are rare, but notable because of their elucidating of the nature of the Y chromosome.
More than two Y chromosomes
Greater degrees of Y chromosome polysomy (having more than one extra copy of the Y chromosome in every cell, e.g., XYYY) are rare. The extra genetic material in these cases can lead to skeletal abnormalities, decreased IQ, and delayed development, but the severity features of these conditions are variable.
XX male syndrome
XX male syndrome occurs when there has been a recombination in the formation of the male gametes, causing the SRY portion of the Y chromosome to move to the X chromosome. When such an X chromosome contributes to the child, the development will lead to a male, because of the SRY gene.
Genetic genealogy
In human genetic genealogy (the application of genetics to traditional genealogy), use of the information contained in the Y chromosome is of particular interest because, unlike other chromosomes, the Y chromosome is passed exclusively from father to son, on the patrilineal line. Mitochondrial DNA, maternally inherited to both sons and daughters, is used in an analogous way to trace the matrilineal line.
Brain function
Research is currently investigating whether male-pattern neural development is a direct consequence of Y-chromosome-related gene expression or an indirect result of Y-chromosome-related androgenic hormone production.[63]
Microchimerism
The presence of male chromosomes in fetal cells in the blood circulation of women was discovered in 1974.[64]
In 1996, it was found that male fetal progenitor cells could persist postpartum in the maternal blood stream for as long as 27 years.[65]
A 2004 study at the Fred Hutchinson Cancer Research Center, Seattle, investigated the origin of male chromosomes found in the peripheral blood of women who had not had male progeny. A total of 120 subjects (women who had never had sons) were investigated, and it was found that 21% of them had male DNA. The subjects were categorised into four groups based on their case histories:[66]
- Group A (8%) had had only female progeny.
- Patients in Group B (22%) had a history of one or more miscarriages.
- Patients Group C (57%) had their pregnancies medically terminated.
- Group D (10%) had never been pregnant before.
The study noted that 10% of the women had never been pregnant before, raising the question of where the Y chromosomes in their blood could have come from. The study suggests that possible reasons for occurrence of male chromosome microchimerism could be one of the following:[66]
- miscarriages,
- pregnancies,
- vanished male twin,
- possibly from sexual intercourse.
A 2012 study at the same institute has detected cells with the Y chromosome in multiple areas of the brains of deceased women.[67]
Cytogenetic band

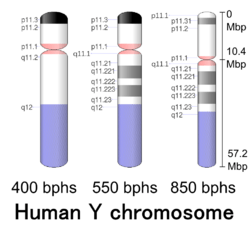
| Chr. | Arm[72] | Band[73] | ISCN start[74] |
ISCN stop[74] |
Basepair start |
Basepair stop |
Stain[75] | Density |
|---|---|---|---|---|---|---|---|---|
| Y | p | 11.32 | 0 | 149 | 1 | 300,000 | gneg | |
| Y | p | 11.31 | 149 | 298 | 300,001 | 600,000 | gpos | 50 |
| Y | p | 11.2 | 298 | 1043 | 600,001 | 10,300,000 | gneg | |
| Y | p | 11.1 | 1043 | 1117 | 10,300,001 | 10,400,000 | acen | |
| Y | q | 11.1 | 1117 | 1266 | 10,400,001 | 10,600,000 | acen | |
| Y | q | 11.21 | 1266 | 1397 | 10,600,001 | 12,400,000 | gneg | |
| Y | q | 11.221 | 1397 | 1713 | 12,400,001 | 17,100,000 | gpos | 50 |
| Y | q | 11.222 | 1713 | 1881 | 17,100,001 | 19,600,000 | gneg | |
| Y | q | 11.223 | 1881 | 2160 | 19,600,001 | 23,800,000 | gpos | 50 |
| Y | q | 11.23 | 2160 | 2346 | 23,800,001 | 26,600,000 | gneg | |
| Y | q | 12 | 2346 | 3650 | 26,600,001 | 57,227,415 | gvar |
See also
- Genealogical DNA test
- Genetic genealogy
- Haplodiploid sex-determination system
- Human Y chromosome DNA haplogroups
- List of Y-STR markers
- Muller's ratchet
- Single nucleotide polymorphism
- Y chromosome Short Tandem Repeat (STR)
- Y linkage
- Y-chromosomal Aaron
- Y-chromosomal Adam
- Y-chromosome haplogroups in populations of the world
References
- "Human Genome Assembly GRCh38 - Genome Reference Consortium". National Center for Biotechnology Information. 2013-12-24. Retrieved 2017-03-04.
- "Homo sapiens Y chromosome genes". CCDS Release 20 for Homo sapiens. 2016-09-08. Retrieved 2017-05-28.
- Strachan T, Read A (2 April 2010). Human Molecular Genetics. Garland Science. p. 45. ISBN 978-1-136-84407-2.
- Genome Decoration Page, NCBI. Ideogram data for Homo sapience (850 bphs, Assembly GRCh38.p3). Last update 2014-06-03. Retrieved 2017-04-26.
- "Ensembl Human MapView release 43". February 2014. Retrieved 2007-04-14.
- Wade N (January 13, 2010). "Male Chromosome May Evolve Fastest". New York Times.
- David Bainbridge, The X in Sex: How the X Chromosome Controls Our Lives, pages 3-5, 13, Harvard University Press, 2003 ISBN 0674016211.
- James Schwartz, In Pursuit of the Gene: From Darwin to DNA, pages 170-172, Harvard University Press, 2009 ISBN 0674034910
- David Bainbridge, The X in Sex: How the X Chromosome Controls Our Lives, pages 65-66, Harvard University Press, 2003 ISBN 0674016211
- Muller HJ (1914). "A gene for the fourth chromosome of Drosophila". Journal of Experimental Zoology. 17 (3): 325–336. doi:10.1002/jez.1400170303.
- Lahn BT, Page DC (October 1999). "Four evolutionary strata on the human X chromosome". Science. 286 (5441): 964–7. doi:10.1126/science.286.5441.964. PMID 10542153.
- Graves JA (March 2006). "Sex chromosome specialization and degeneration in mammals". Cell. 124 (5): 901–14. doi:10.1016/j.cell.2006.02.024. PMID 16530039.
- Graves JA, Koina E, Sankovic N (June 2006). "How the gene content of human sex chromosomes evolved". Current Opinion in Genetics & Development. 16 (3): 219–24. doi:10.1016/j.gde.2006.04.007. PMID 16650758.
- Bachtrog D (February 2013). "Y-chromosome evolution: emerging insights into processes of Y-chromosome degeneration". Nature Reviews. Genetics. 14 (2): 113–24. doi:10.1038/nrg3366. PMC 4120474. PMID 23329112.
- Hamilton J (January 13, 2010). "Human Male: Still A Work in Progress". NPR.
- Warren WC, Hillier LW, Marshall Graves JA, Birney E, Ponting CP, Grützner F, et al. (May 2008). "Genome analysis of the platypus reveals unique signatures of evolution". Nature. 453 (7192): 175–83. Bibcode:2008Natur.453..175W. doi:10.1038/nature06936. PMC 2803040. PMID 18464734.
- Veyrunes F, Waters PD, Miethke P, Rens W, McMillan D, Alsop AE, et al. (June 2008). "Bird-like sex chromosomes of platypus imply recent origin of mammal sex chromosomes". Genome Research. 18 (6): 965–73. doi:10.1101/gr.7101908. PMC 2413164. PMID 18463302.
- Grützner F, Rens W, Tsend-Ayush E, El-Mogharbel N, O'Brien PC, Jones RC, et al. (December 2004). "In the platypus a meiotic chain of ten sex chromosomes shares genes with the bird Z and mammal X chromosomes". Nature. 432 (7019): 913–7. Bibcode:2004Natur.432..913G. doi:10.1038/nature03021. PMID 15502814.
- Watson JM, Riggs A, Graves JA (October 1992). "Gene mapping studies confirm the homology between the platypus X and echidna X1 chromosomes and identify a conserved ancestral monotreme X chromosome". Chromosoma. 101 (10): 596–601. doi:10.1007/BF00360536. PMID 1424984.
- Graves JA (2004). "The degenerate Y chromosome--can conversion save it?". Reproduction, Fertility, and Development. 16 (5): 527–34. doi:10.1071/RD03096. PMID 15367368.
- Goto H, Peng L, Makova KD (February 2009). "Evolution of X-degenerate Y chromosome genes in greater apes: conservation of gene content in human and gorilla, but not chimpanzee". Journal of Molecular Evolution. 68 (2): 134–44. Bibcode:2009JMolE..68..134G. doi:10.1007/s00239-008-9189-y. PMID 19142680.
- Hughes JF, Skaletsky H, Pyntikova T, Minx PJ, Graves T, Rozen S, et al. (September 2005). "Conservation of Y-linked genes during human evolution revealed by comparative sequencing in chimpanzee". Nature. 437 (7055): 100–3. Bibcode:2005Natur.437..100H. doi:10.1038/nature04101. PMID 16136134.
- Hsu C. "Biologists Debunk the 'Rotting' Y Chromosome Theory, Men Will Still Exist". Medical Daily.
- Lindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, Kamal M, et al. (December 2005). "Genome sequence, comparative analysis and haplotype structure of the domestic dog". Nature. 438 (7069): 803–19. Bibcode:2005Natur.438..803L. doi:10.1038/nature04338. PMID 16341006.
- Brandeis M (May 2018). "New-age ideas about age-old sex: separating meiosis from mating could solve a century-old conundrum". Biological Reviews of the Cambridge Philosophical Society. 93 (2): 801–810. doi:10.1111/brv.12367. PMID 28913952.
- Bernstein H, Hopf FA, Michod RE (1987). "The molecular basis of the evolution of sex". Molecular Genetics of Development. Advances in Genetics. 24. pp. 323–70. doi:10.1016/S0065-2660(08)60012-7. ISBN 9780120176243. PMID 3324702.
- Liu Z, Venkatesh SS, Maley CC (October 2008). "Sequence space coverage, entropy of genomes and the potential to detect non-human DNA in human samples". BMC Genomics. 9 (1): 509. doi:10.1186/1471-2164-9-509. PMC 2628393. PMID 18973670. Fig. 6, using the Lempel-Ziv estimators of entropy rate.
- Charlesworth B, Charlesworth D (November 2000). "The degeneration of Y chromosomes". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 355 (1403): 1563–72. doi:10.1098/rstb.2000.0717. PMC 1692900. PMID 11127901.
- Rozen S, Skaletsky H, Marszalek JD, Minx PJ, Cordum HS, Waterston RH, et al. (June 2003). "Abundant gene conversion between arms of palindromes in human and ape Y chromosomes". Nature. 423 (6942): 873–6. Bibcode:2003Natur.423..873R. doi:10.1038/nature01723. PMID 12815433.
- Marchal JA, Acosta MJ, Bullejos M, Díaz de la Guardia R, Sánchez A (2003). "Sex chromosomes, sex determination, and sex-linked sequences in Microtidae". Cytogenetic and Genome Research. 101 (3–4): 266–73. doi:10.1159/000074347. PMID 14684993.
- Wilson MA, Makova KD (2009). "Genomic analyses of sex chromosome evolution". Annual Review of Genomics and Human Genetics. 10 (1): 333–54. doi:10.1146/annurev-genom-082908-150105. PMID 19630566.
- Just W, Baumstark A, Süss A, Graphodatsky A, Rens W, Schäfer N, et al. (2007). "Ellobius lutescens: sex determination and sex chromosome". Sexual Development. 1 (4): 211–21. doi:10.1159/000104771. PMID 18391532.
- Arakawa Y, Nishida-Umehara C, Matsuda Y, Sutou S, Suzuki H (2002). "X-chromosomal localization of mammalian Y-linked genes in two XO species of the Ryukyu spiny rat". Cytogenetic and Genome Research. 99 (1–4): 303–9. doi:10.1159/000071608. PMID 12900579.
- Hoekstra HE, Edwards SV (September 2000). "Multiple origins of XY female mice (genus Akodon): phylogenetic and chromosomal evidence". Proceedings. Biological Sciences. 267 (1455): 1825–31. doi:10.1098/rspb.2000.1217. PMC 1690748. PMID 11052532.
- Ortiz MI, Pinna-Senn E, Dalmasso G, Lisanti JA (2009). "Chromosomal aspects and inheritance of the XY female condition in Akodon azarae (Rodentia, Sigmodontinae)". Mammalian Biology. 74 (2): 125–129. doi:10.1016/j.mambio.2008.03.001.
- Charlesworth B, Dempsey ND (April 2001). "A model of the evolution of the unusual sex chromosome system of Microtus oregoni". Heredity. 86 (Pt 4): 387–94. doi:10.1046/j.1365-2540.2001.00803.x. PMID 11520338.
- Zhou Q, Wang J, Huang L, Nie W, Wang J, Liu Y, et al. (2008). "Neo-sex chromosomes in the black muntjac recapitulate incipient evolution of mammalian sex chromosomes". Genome Biology. 9 (6): R98. doi:10.1186/gb-2008-9-6-r98. PMC 2481430. PMID 18554412.
- Hamilton WD (April 1967). "Extraordinary sex ratios. A sex-ratio theory for sex linkage and inbreeding has new implications in cytogenetics and entomology". Science. 156 (3774): 477–88. Bibcode:1967Sci...156..477H. doi:10.1126/science.156.3774.477. PMID 6021675.
- Schartl M (July 2004). "A comparative view on sex determination in medaka". Mechanisms of Development. 121 (7–8): 639–45. doi:10.1016/j.mod.2004.03.001. PMID 15210173.
- Cortez D, Marin R, Toledo-Flores D, Froidevaux L, Liechti A, Waters PD, et al. (April 2014). "Origins and functional evolution of Y chromosomes across mammals". Nature. 508 (7497): 488–93. Bibcode:2014Natur.508..488C. doi:10.1038/nature13151. PMID 24759410.
- Deakin JE, Graves JA, Rens W (2012). "The evolution of marsupial and monotreme chromosomes". Cytogenetic and Genome Research. 137 (2–4): 113–29. doi:10.1159/000339433. PMID 22777195.
- National Library of Medicine's Genetic Home Reference
- "Definition of holandric | Dictionary.com". www.dictionary.com. Retrieved 2020-01-21.
- Forsberg LA (May 2017). "Loss of chromosome Y (LOY) in blood cells is associated with increased risk for disease and mortality in aging men". Human Genetics. 136 (5): 657–663. doi:10.1007/s00439-017-1799-2. PMC 5418310. PMID 28424864.
- Forsberg LA, Rasi C, Malmqvist N, Davies H, Pasupulati S, Pakalapati G, et al. (June 2014). "Mosaic loss of chromosome Y in peripheral blood is associated with shorter survival and higher risk of cancer". Nature Genetics. 46 (6): 624–8. doi:10.1038/ng.2966. PMC 5536222. PMID 24777449.
- Coghlan A (13 December 2014). "Y men are more likely to get cancer than women". New Scientist: 17.
- Dumanski JP, Rasi C, Lönn M, Davies H, Ingelsson M, Giedraitis V, et al. (January 2015). "Mutagenesis. Smoking is associated with mosaic loss of chromosome Y". Science. 347 (6217): 81–3. Bibcode:2015Sci...347...81D. doi:10.1126/science.1262092. PMC 4356728. PMID 25477213.
- Science Daily, Apr. 3, 2008.
- Pertea M, Salzberg SL (2010). "Between a chicken and a grape: estimating the number of human genes". Genome Biology. 11 (5): 206. doi:10.1186/gb-2010-11-5-206. PMC 2898077. PMID 20441615.
- "Statistics & Downloads for chromosome Y". HUGO Gene Nomenclature Committee. 2017-05-12. Retrieved 2017-05-19.
- "Chromosome Y: Chromosome summary - Homo sapiens". Ensembl Release 88. 2017-03-29. Retrieved 2017-05-19.
- "Human chromosome Y: entries, gene names and cross-references to MIM". UniProt. 2018-02-28. Retrieved 2018-03-16.
- "Homo sapiens Y chromosome coding genes". National Center for Biotechnology Information Gene Database. 2017-05-19. Retrieved 2017-05-20.
- "Homo sapiens Y chromosome non-coding genes". 2017-05-19. Retrieved 2017-05-20.
- "Homo sapiens Y chromosome non-coding pseudo genes". 2017-05-19. Retrieved 2017-05-20.
- Veerappa AM, Padakannaya P, Ramachandra NB (August 2013). "Copy number variation-based polymorphism in a new pseudoautosomal region 3 (PAR3) of a human X-chromosome-transposed region (XTR) in the Y chromosome". Functional & Integrative Genomics. 13 (3): 285–93. doi:10.1007/s10142-013-0323-6. PMID 23708688.
- Raudsepp T, Chowdhary BP (6 January 2016). "The Eutherian Pseudoautosomal Region". Cytogenetic and Genome Research. 147 (2–3): 81–94. doi:10.1159/000443157. PMID 26730606.
- 1950-, Nussbaum, Robert L. (2007). Thompson & Thompson genetics in medicine. McInnes, Roderick R., Willard, Huntington F., Hamosh, Ada., Thompson, Margaret W. (Margaret Wilson), 1920- (7th. ed.). Philadelphia: Saunders/Elsevier. ISBN 978-1416030805. OCLC 72774424.CS1 maint: numeric names: authors list (link)
- Jacobs PA, Brunton M, Melville MM, Brittain RP, McClemont WF (December 1965). "Aggressive behavior, mental sub-normality and the XYY male". Nature. 208 (5017): 1351–2. Bibcode:1965Natur.208.1351J. doi:10.1038/2081351a0. PMID 5870205.
- Richardson SS (2013). Sex Itself: The Search for Male & Female in the Human Genome. Chicago: U. of Chicago Press. p. 84. ISBN 978-0-226-08468-8.
- Witkin HA, Mednick SA, Schulsinger F, Bakkestrom E, Christiansen KO, Goodenough DR, et al. (August 1976). "Criminality in XYY and XXY men". Science. 193 (4253): 547–55. Bibcode:1976Sci...193..547W. doi:10.1126/science.959813. PMID 959813.
- Witkin HA, Goodenough DR, Hirschhorn K (1977). "XYY Men: Are They Criminally Aggressive?". The Sciences. 17 (6): 10–13. doi:10.1002/j.2326-1951.1977.tb01570.x. PMID 11662398.
- Kopsida E, Stergiakouli E, Lynn PM, Wilkinson LS, Davies W (2009). "The Role of the Y Chromosome in Brain Function" (PDF). Open Neuroendocrinology Journal. 2: 20–30. doi:10.2174/1876528900902010020. PMC 2854822. PMID 20396406.
- Schröder J, Thlikainen A, de la Chapelle A (1974). "Fetal leukocytes in the maternal circulation after delivery: Cytological aspects". Transplantation. 17 (4): 346–354. doi:10.1097/00007890-197404000-00003. ISSN 0041-1337.
- Bianchi DW, Zickwolf GK, Weil GJ, Sylvester S, DeMaria MA (January 1996). "Male fetal progenitor cells persist in maternal blood for as long as 27 years postpartum". Proceedings of the National Academy of Sciences of the United States of America. 93 (2): 705–8. Bibcode:1996PNAS...93..705B. doi:10.1073/pnas.93.2.705. PMC 40117. PMID 8570620.
- Yan Z, Lambert NC, Guthrie KA, Porter AJ, Loubiere LS, Madeleine MM, et al. (August 2005). "Male microchimerism in women without sons: quantitative assessment and correlation with pregnancy history" (full text). The American Journal of Medicine. 118 (8): 899–906. doi:10.1016/j.amjmed.2005.03.037. PMID 16084184.
- Chan WF, Gurnot C, Montine TJ, Sonnen JA, Guthrie KA, Nelson JL (26 September 2012). "Male microchimerism in the human female brain". PLOS One. 7 (9): e45592. Bibcode:2012PLoSO...745592C. doi:10.1371/journal.pone.0045592. PMC 3458919. PMID 23049819.
- Genome Decoration Page, NCBI. Ideogram data for Homo sapience (400 bphs, Assembly GRCh38.p3). Last update 2014-03-04. Retrieved 2017-04-26.
- Genome Decoration Page, NCBI. Ideogram data for Homo sapience (550 bphs, Assembly GRCh38.p3). Last update 2015-08-11. Retrieved 2017-04-26.
- International Standing Committee on Human Cytogenetic Nomenclature (2013). ISCN 2013: An International System for Human Cytogenetic Nomenclature (2013). Karger Medical and Scientific Publishers. ISBN 978-3-318-02253-7.
- Sethakulvichai W, Manitpornsut S, Wiboonrat M, Lilakiatsakun W, Assawamakin A, Tongsima S (2012). Estimation of band level resolutions of human chromosome images. In Computer Science and Software Engineering (JCSSE), 2012 International Joint Conference on. pp. 276–282. doi:10.1109/JCSSE.2012.6261965. ISBN 978-1-4673-1921-8.
- "p": Short arm; "q": Long arm.
- For cytogenetic banding nomenclature, see article locus.
- These values (ISCN start/stop) are based on the length of bands/ideograms from the ISCN book, An International System for Human Cytogenetic Nomenclature (2013). Arbitrary unit.
- gpos: Region which is positively stained by G banding, generally AT-rich and gene poor; gneg: Region which is negatively stained by G banding, generally CG-rich and gene rich; acen Centromere. var: Variable region; stalk: Stalk.
External links
| Wikimedia Commons has media related to Y chromosomes. |
- Genetic Genealogy: About the use of mtDNA and Y chromosome analysis in ancestry testing
- Ensembl genome browser
- http://www.ncbi.nlm.nih.gov/mapview/maps.cgi?taxid=9606&chr=Y
- Human Genome Project Information—Human Chromosome Y Launchpad
- On Topic: Y Chromosome—From the Whitehead Institute for Biomedical Research
- Nature—focus on the Y chromosome
- National Human Genome Research Institute (NHGRI)—Use of Novel Mechanism Preserves Y chromosome Genes
- Ysearch.org – Public Y-DNA database
- Y chromosome Consortium (YCC)
- NPR's Human Male: Still A Work In Progress
| Biological terms | |
|---|---|
| Sexual reproduction |
|
| Sexuality | |
| |