Gorgonopsia
Gorgonopsia is an extinct clade of non-mammalian synapsids from the Permian period. Gorgonopsians were quadrupedal predators with prominent canine teeth, and the largest species were the apex predators of their ecosystems.[1] Like other non-mammalian synapsids, gorgonopsians were once described as "mammal-like reptiles", due to their mix of mammalian and reptilian traits. However, this description is no longer considered accurate as they are not reptiles.[2] Rather, as therapsids, they are closely related to mammals. Gorgonopsian fossils have been found in Russia and Africa.[3]
| Gorgonopsians | |
|---|---|
 | |
| Skeleton of Sauroctonus parringtoni | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Therapsida |
| Clade: | Theriodontia |
| Suborder: | †Gorgonopsia Seeley, 1895 |
| Family: | †Gorgonopsidae Lydekker, 1890 |
| Genera | |
Description
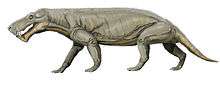
Gorgonopsians are the most morphologically homogeneous of the major therapsid clades.[2] A distinctive feature of gorgonopsians is the large size of their canine teeth, which has led them to be compared to saber-toothed cats, though they have few other similarities. In the majority of gorgonopsians, only the incisors and canines were large; the postcanine teeth were reduced in both size and number, to the point where one genus, Clelandina, had no postcanine teeth at all.
The dentition of Gorgonopsia was intermediate between that of earlier amniotes and that of mammals. It was strongly heterodont, with distinct incisors, canines, and postcanine teeth; the postcanine teeth are homologous to the premolars and molars of mammals.[4] Unlike mammals, however, gorgonopsians were polyphyodont, with teeth replaced multiple times throughout their lifetime.[5] The postcanine teeth were replaced more slowly than the other teeth, likely due to their lack of functional significance.[4] In gorgonopsians, as in some other therapsids, the upper canines had an unusual pattern of replacement. There were two canine alveoli in each maxilla, but only one functional tooth at a time on each side; one alveolus would bear a functional tooth while the other was growing a new one.[5] The left and right sides were not in sync, so the functional canine could be in the anterior alveolus on one side and the posterior alveolus on the other.
The gorgonopsians' mammalian specializations include differentiated (heterodont) tooth shape, a fully developed temporal fenestra, pillar-like rear legs, a vaulted palate that may have facilitated breathing while holding prey, and incipiently developed ear bones.[6] Despite their conservative anatomy, gorgonopsians varied considerably in size.[2] The smallest gorgonopsians had skulls less than 15 centimetres (5.9 in) long, whereas the largest were among the largest carnivores of the late Permian. The largest species known, Inostrancevia, was the size of a large bear with a 45-centimetre-long (18 in) skull, and 12-centimetre-long (4.7 in) sabre-like teeth. The integument of gorgonopsians is unknown, but they are thought to have lacked a full coat of fur.[7] Like most therapsids, they are assumed to have been terrestrial, which is supported both by their morphology and bone microanatomy.[8]
Evolutionary history
Gorgonopsians are theriodonts, a major group of therapsids that included the ancestors of mammals. They evolved in the Middle Permian, from a reptile-like therapsid that also lived in that period. The early gorgonopsians were small, being no larger than a dog. The extinction of dinocephalians (which dominated the Middle Permian world) led to gorgonopsids becoming the apex predators of the Late Permian. Some had approached the size of a rhinoceros, such as Inostrancevia, the largest gorgonopsian. A nearly complete fossil of Rubidgea has been found in South Africa.[9] The Gorgonopsia became extinct at the end of the Permian period, being the only theriodont line to be terminated by this mass extinction.
Classification
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogeny of Gorgonopsia according to Bendel et al. 2018[4] |
Gorgonopsians constitute one of the six major subclades of Therapsida.[2] As therapsids, they are part of the mammalian stem group. Within Therapsida, they are often classified alongside therocephalians and cynodonts as members of Theriodontia, but this is debated.[2]
The taxonomy and internal phylogenetic relationships of Gorgonopsia are poorly understood, and they have been considered the most poorly-studied clade of therapsids.[2] Many species of gorgonopsian were named by Robert Broom, a notorious splitter, many of whose taxa are now regarded as invalid.[10] Many species distinctions among gorgonopsians have been based on minor differences which can be easily affected by preservation.[1] As a result of these difficulties, relatively little research has gone into resolving gorgonopsian taxonomy and phylogeny.[1]
Many species once considered to have been basal gorgonopsians are now classified as members of Biarmosuchia.[2] The remaining gorgonopsians have traditionally been assigned to the single family Gorgonopidae.[1]
The most well-defined subgroup within Gorgonopsia is Rubidgeinae, which comprises several genera of large, robust gorgonopsians from Africa.[1] Rubidgeinae is within a larger clade of gorgonopsians endemic to Africa.[3] Historically, several subfamilies of gorgonopsian were recognized, especially Gorgonopsinae, Rubidgeinae and Inostranceviinae. However, only Rubidgeinae is currently recognized, pending further research.[1]
The most comprehensive review of the group is by Sigogneau-Russell, 1989.[11] However, many gorgonopsian taxa have had their taxonomy changed since then.[1] In all, there are 25 genera and 41 species, with the genera described most completely being Dinogorgon, Inostrancevia and Rubidgea.


- Order Therapsida
- SUBORDER GORGONOPSIA
- Family Gorgonopsidae
- Aelurosaurus
- Aloposaurus
- Arctognathus
- Arctops
- Broomisaurus
- Cerdorhinus
- Cynariops
- Cyonosaurus
- Eriphostoma
- Kamagorgon
- Lycaenops
- "Njalila"
- Nochnitsa
- Paragalerhinus
- Smilesaurus
- Suchogorgon
- Viatkogorgon
- "Gorgonops" dixeyi
- Subfamily Gorgonopsinae
- Subfamily Inostranceviinae
- Subfamily Rubidgeinae
- Family Gorgonopsidae
Paleobiology
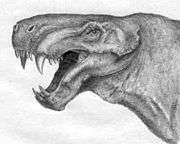
The habitual head posture was anteriorly tilted revealed by the orientation of the horizontal semicircular canal.[12] They possessed relatively large floccular fossa, a portion of the cerebellum that controls gaze stability.[12]
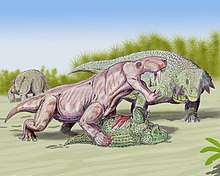
Gorgonopsians were likely active predators, based on their powerful senses of smell and their fairly good eyesight. Gorgonopsians were also seemingly plantigrade walkers, based on their skeletal morphology, and probably possessed a gait similar to a "crocodilian high-walk". This posture would have permitted them to be much faster than many of their potential prey species, such as dicynodonts and pareiasaurs, particularly in conjunction with their reduced, almost mammalian phalangeal formula and more symmetrical feet. Their hunting strategy was probably one of ambush; lying in wait before lunging at speed to grapple prey with their front limbs and attack their prey with their saber-teeth. Unlike later sabertooths such as the machairodontines, gorgonopsians likely were less precise in regards to bite placement; as they had reptilian jaws and tooth arrangements, gorgonopsians probably hunted by using a bite-and-retreat technique to weaken and debilitate their victim before moving in to attack the throat, underbelly and other vulnerable areas. Due to the fact that most gorgonopsians lack any post-canine cutting teeth, meat would have been torn away from a carcass using the powerful jaw muscles and incisors before being gulped down and swallowed whole.[13]:204–207
During most of the late Permian, gorgonopsians were the apex predators of their ecosystems. However, earlier in their evolutionary history, during the middle Permian, gorgonopsians such as the recently discovered Nochnitsa were fairly small, rare animals.[3] The discovery of Nochnitsa in conjunction with the larger therocephalian Gorynychus indicates that after the extinction of dinocephalians but before gorgonopsians rose to dominance, basal therocephalians were the apex predators of Permian ecosystems.[14][15]
Some gorgonopsians, such as the small, basal gorgonopsian Viatkogorgon, were nocturnal.[3] Others, such as the large rubidgeine Clelandina, were diurnal.[1]
Paleopathology
A fossil lower arm bone described by Kyle Keto et al. in 2020 indicates that gorgonopsids were capable of rapid healing, much like the unrelated dinosaurs and many modern animals are. The specimen, NHCC LB396, contains bone lesions that are the result of an inflammation of the connective tissue around the periosteum that was overlain by bone growth created as the gorgonopsid healed. The injury that caused the bone growth is unclear, but the lesions resulting from healing this injury are believed to have developed within one year of the animal's life.[16]
References
- Kammerer, Christian F. (2016). "Systematics of the Rubidgeinae (Therapsida: Gorgonopsia)". PeerJ. 4: e1608. doi:10.7717/peerj.1608. PMC 4730894. PMID 26823998.
- Angielczyk, Kenneth D.; Kammerer, Christian F. (2018). "Non-Mammalian synapsids: the deep roots of the mammalian family tree". In Zachos, Frank E.; Asher, Robert J. (eds.). Mammalian Evolution, Diversity and Systematics. Berlin: De Gruyter. ISBN 9783110275902.
- Kammerer, Christian F.; Masyutin, Vladimir (2018). "Gorgonopsian therapsids (Nochnitsa gen. nov. and Viatkogorgon) from the Permian Kotelnich locality of Russia". PeerJ. 6: e4954. doi:10.7717/peerj.4954. PMC 5995105. PMID 29900078.
- Bendel, Eva-Maria; Kammerer, Christian F.; Kardjilov, Nikolay; Fernandez, Vincent; Fröbisch, Jörg (2018). "Cranial anatomy of the gorgonopsian Cynariops robustus based on CT-reconstruction". PLoS ONE. 13 (11): e0207367. doi:10.1371/journal.pone.0207367. PMC 6261584. PMID 30485338.
- Kermack, K. A. (1956). "Tooth replacement in mammal-like reptiles of the suborders Gorgonopsia and Therocephalia". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 240: 95–133. doi:10.1098/rstb.1956.0013.
- Laurin, Michel (1998). "New data on the cranial anatomy of Lycaenops (Synapsida, Gorgonopsidae), and reflections on the possible presence of streptostyly in gorgonopsians". Journal of Vertebrate Paleontology. 18 (4): 765–776. doi:10.1080/02724634.1998.10011105. JSTOR 4523954.
- Ruben, John A.; Jones, Terry D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". American Zoologist. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- Kriloff, A.; Germain, D.; Canoville, A.; Vincent, P.; Sache, M.; Laurin, M. (2008). "Evolution of bone microanatomy of the tetrapod tibia and its use in palaeobiological inference". Journal of Evolutionary Biology. 21 (3): 807–826. doi:10.1111/j.1420-9101.2008.01512.x. PMID 18312321.
- University of Washington (10 December 1998). "First Complete Fossil Of Fierce Prehistoric Predator Found In South Africa". ScienceDaily.
- Wyllie, Alistair (2003). "A review of Robert Broom's therapsid holotypes: have they survived the test of time?". Palaeontologia Africana. 39: 1–19. hdl:10539/15952.
- Sigogneau-Russell, Denise (1989). Wellnhofer, Peter (ed.). Theriodontia I: Phthinosuchia, Biarmosuchia, Eotitanosuchia, Gorgonopsia. Encyclopedia of Paleoherpetology. 17 B/I. Stuttgart: Gustav Fischer Verlag. ISBN 978-3437304873.
- Araújo, Ricardo; Fernandez, Vincent; Polcyn, Michael J.; Fröbisch, Jörg; Martins, Rui M. S. (2017). "Aspects of gorgonopsian paleobiology and evolution: insights from the basicranium, occiput, osseous labyrinth, vasculature, and neuroanatomy". PeerJ. 5: e3119. doi:10.7717/peerj.3119. PMC 5390774. PMID 28413721.
- Antón, Mauricio (2013). Sabertooth. Bloomington, Indiana: University of Indiana Press. ISBN 9780253010421.
- North Carolina Museum of Natural Sciences (8 June 2018). "'Monstrous' new Russian saber-tooth fossils clarify early evolution of mammal lineage". ScienceDaily.
- Kammerer, Christian F.; Masyutin, Vladimir (2018). "A new therocephalian (Gorynychus masyutinae gen. et sp. nov.) from the Permian Kotelnich locality, Kirov Region, Russia". PeerJ. 6: e4933. doi:10.7717/peerj.4933. PMC 5995100. PMID 29900076.
- https://blogs.scientificamerican.com/laelaps/sabertoothed-protomammal-was-a-quick-healer/
Further reading
- Bakker, R.T. (1986), The Dinosaur Heresies, Kensington Publishing Corp.
- Cox, B. and Savage, R.J.G. and Gardiner, B. and Harrison, C. and Palmer, D. (1988) The Marshall illustrated encyclopedia of dinosaurs & prehistoric animals, 2nd Edition, Marshall Publishing.
- Fenton, C.L.and Fenton, M.A. (1958) The Fossil Book, Doubleday Publishing.
- Hore, P.M. (2005), The Dinosaur Age, Issue #18. National Dinosaur Museum.
- Ward, P.D. (2004), Gorgon, Viking Penguin.
| |||||||||
| Basal Gorgonopsians |  | ||||||||
| Gorgonopsidae |
| ||||||||
| Related categories |
| ||||||||