Tick-borne encephalitis virus
Tick-borne encephalitis virus (TBEV) is a virus associated with tick-borne encephalitis.
| Tick-borne encephalitis virus | |
|---|---|
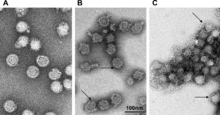 | |
| TBEV at different pH levels | |
| Virus classification | |
| (unranked): | Virus |
| Realm: | Riboviria |
| Kingdom: | Orthornavirae |
| Phylum: | Kitrinoviricota |
| Class: | Flasuviricetes |
| Order: | Amarillovirales |
| Family: | Flaviviridae |
| Genus: | Flavivirus |
| Species: | Tick-borne encephalitis virus |
Taxonomy
TBEV is a member of the genus Flavivirus. Other close relatives, members of the TBEV serocomplex, include Omsk hemorrhagic fever virus, Kyasanur Forest disease virus, Alkhurma virus, Louping ill virus and Langat virus.[1]
Subtypes
TBEV has three subtypes:
- Western European subtype (formerly Central European encephalitis virus, CEEV; principal tick vector: Ixodes ricinus);
- Siberian subtype (formerly West Siberian virus; principal tick vector: Ixodes persulcatus);
- Far Eastern subtype (formerly Russian Spring Summer encephalitis virus, RSSEV; principal tick vector: Ixodes persulcatus).[2]
The reference strain is the Sofjin strain.[3]
Virology
TBEV is a positive-sense single stranded RNA virus, contained in a 40-60 nm spherical, enveloped capsid.[4] The TBEV genome is approximately 11kb in size, which contains a 5' cap, a single open reading frame with 3' and 5' UTRs, and is without polyadenylation.[4] Like other flaviviruses,[5] the TBEV genome codes for ten viral proteins, three structural, and seven nonstructural (NS). The structural proteins are C (capsid), PrM (premembrane, which is cleaved to produce the final membrane protein, M), and E (envelope). The seven nonstructural proteins are: NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5. The role of some nonstructural proteins is known, NS5 serves as RNA-dependent polymerase, NS3 has protease (in complex with NS2B) and helicase activity.[6][4] Structural and nonstructral proteins are not required for the genome to be infectious.[4] All viral proteins are expressed as a single large polyprotein, with the order C, PrM, E, NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5.[6]
Life cycle
Vector
Infection of the vector begins when a tick takes a blood meal from an infected host. This can occur at any part of the tick's life cycle but a "horizontal" transmission between infected nymphs and uninfected larvae co-feeding on the same host is thought to be key in maintaining the circulation of TBEV.[7][4] TBEV in the blood of the host infects the tick through the midgut, from where it can pass to the salivary glands to be passed to the next host. In non-adult ticks, TBEV is transmitted transtadially by infecting cells that are not destroyed during molting, thus the tick remains infectious throughout its life.[7] Infected adult ticks may be able to lay eggs that are infected, transmitting the virus transorvarially.[8]
Viral
In humans, the infection begins in the skin (with the exception of foodborne cases, about 1% of infections) at the site of the bite of an infected tick, where Langerhans cells and macrophages in the skin are preferentially targeted.[6] TBEV envelope (E) proteins recognize heparan sulfate (and likely other receptors) on the host cell surface and are endocytosed via the clathrin mediated pathway. Acidification of the late endosome triggers a conformational change in the E proteins, resulting in fusion, followed by uncoating, and release of the single-stranded RNA genome into the cytoplasm.[9][4] The viral polyprotein is translated and inserts into the ER membrane, where it is processed on the cytosolic side by host peptidases and in the lumen by viral enzyme action. The viral proteins C, NS3, and NS5 are cleaved into the cytosol (though NS3 can complex with NS2B or NS4A to perform proteolytic or helicase activity), while the remaining nonstructural proteins alter the structure of the ER membrane. This altered membrane permits the assembly of replication complexes, where the viral genome is replicated by the viral RNA-dependent polymerase, NS5.[9][6] Newly replicated viral RNA genomes are then packaged by the C proteins while on the cytosolic side of the of the ER memebrane, forming the immature nucleocapsid, and gain E and PrM proteins, arranged as a heterodimer, during budding into the lumen of the ER. The immature virion is spiky and geometric in comparison to the mature particle. The particle passes through the golgi apparatus and trans-golgi network, under increasingly acidic conditions, by which the virion matures with cleavage of the Pr segment from the M protein and formation fusion competent E protein homodimers. Though the cleaved Pr segment remains associated with protein complex until exit.[4][9] The virus is released from the host cell upon fusion of the transport vesicle with the host cell membrane, the cleaved Pr now segments dissociate, resulting in a fully mature, infectious virus.[4][9] However, partially mature and immature viruses are sometimes released as well; immature viruses are noninfectious as the E proteins are not fusion competent, partially mature viruses are still capable of infection.[9]
Pathogenesis and immune response
With the exception of food-borne cases, infection begins in the skin at the site of the tick bite. Skin dendritic (or Langerhans) cells (DCs) are preferentially targeted.[6] Initially, the virus replicates locally and immune response is triggered when viral components are recognized by cytosolic pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs).[10] Recognition causes the release of cytokines including interferons (IFN) α, β , and γ and chemokines, attracting migratory immune cells to the site of the bite.[6] The infection may be halted at this stage and cleared, before the onset of noticeable symptoms. Notably, tick saliva enhances infection by modulating host immune response, dampening apoptotic signals.[10] If the infection continues, migratory DCs and macrophages become infected and travel to the local draining lymph node where activation of polymorphonuclear leukocytes, monocytes and the complement system are activated.[10]
The draining lymph node can also serve as a viral amplification site, from where TBEV gains systemic access. This viremic stage corresponds to the first symptomatic phase in the prototypical biphasic pattern of tick-borne encephalitis.[4] TBEV has a strong preference for neuronal tissue, and is neuroinvasive.[11] The initial viremic stage allows access to a number of the preferential tissues. However, the exact mechanism by which TBEV crosses into the central nervous system (CNS) is unclear.[11][10][8][4] There are several proposed mechanism for TBEV breaching the blood-brain barrier (BBB): 1)The "Trojan Horse" mechanism, whereby TBEV gains access to the CNS while infecting an immune cell that passes through the BBB;[10][6][11] 2) Disruption and increased permeability of the BBB by immune immune cytokines;[11] 3) Via infection of the olfactory neurons;[6] 4) Via retrograde transport along peripheral nerves to the CNS;[6] 5) Infection of the cells that make up part of the BBB.[6][10]
CNS infection brings on the second phase in the classic biphasic infection pattern associated with the European subtype. CNS disease is immunopathological; release of inflammatory cytokines coupled with the action of cytotoxic CD8+ T cells and possibly NK cells results in inflammation and apoptosis of infected cells that is responsible for many of the CNS symptoms.[10][11]
Humoral response
TBEV specific IgM and IgG antibodies are produced in response to infection.[4] IgM antibodies appear and peak first, as well as reaching higher levels, and typically dissipates in about 1.5 months post infection, though there exists considerable variation from patient to patient. IgG levels peak at about 6 weeks after the appearance of CNS symptoms, then decline slightly but do not dissipate, likely conferring life long immunity to the patient.[4][6]
Evolution
The ancestor of the extant strains appears to have separated into several clades approximately 2750 years ago.[12] The Siberian and Far Eastern subtypes diverged about 2250 years ago.
A second analysis suggests an earlier date of evolution (3300 years ago) with a rapid increase in the number of strains starting ~300 years ago.[13]
This virus has been transmitted at least three times into Japan between 260–430 years ago.[14][15]
The strains circulating in Latvia appear to have originated from both Russia and Western Europe[16] while those in Estonia appear to have originated in Russia.[17] The Lithuanian strains appear to be related to those from Western Europe.[18]
Phylogenetic analysis indicates that the European and Siberian TBEV sub-types are closely related while the Far-eastern sub-type is closer to the Louping Ill Virus.[1] However, in antigenic relatedness, based on the E, NS3, and NS5 proteins, all three sub-types are highly similar, and Louping Ill virus is the closest relative outside the collective TBEV group.[19]
History
Though the first description of what may have been TBE appears in records in the 1700s in Scandinavia,[11] identification of the TBEV virus occurred in the Soviet Union in the 1930s.[20] The investigation began due to an outbreak of what was believed to be Japanese Encephalitis ("Summer encephalitis"), among Soviet troops stationed along the border with the Japanese empire (present day People's Republic of China), near the Far Eastern city of Khabarovsk. The expedition was lead by virologist Lev A. Zilber, who assembled a team of twenty young scientists in a number of related fields such as acarology, microbiology, neurology, and epidemiology.[21][20] The expedition arrived in Khabarovsk on May 15, 1937, and divided into squads, Northern- led by Elizabeth N. Levkovich and working in the Khabarovski Krai- and Southern- led by Alexandra D. Sheboldaeva, working in the Primorski Krai.[20]
Inside the month of May, the expedition had identified ticks as the likely vector, collected I. persucatus ticks by exposure of bare skin by entomologist Alexander V. Gutsevich, and virologist Mikhail P. Chumakov had isolated the virus from ticks feeding on intentionally infected mice. During the summer, five expeditions members became infected with TBEV, and while there were no fatalities, three of the five suffered damaging sequelae.[20]
The expedition returned in mid August and in October of 1937 Zilber and Sheboldova were arrested, falsely accused of spreading Japanese encephalitis. Expedition epidemiologist Tamara M. Safonov, was arrested the following January for protesting the charges against Zilber and Sheboldova. As a consequence of the arrests, the one of the important initial works was published under the authorship of expedition acarologist, Vasily S. Mironov. Zilber was released in 1939 and managed to restore, along with Sheboldova, co-authorship on this initial work; however, Safanov and Sheboldova (who was not released) spent 18 years in labor camps.[20][21]
References
- Mansfield KL, Johnson N, Phipps LP, Stephenson JR, Fooks AR, Solomon T (August 2009). "Tick-borne encephalitis virus - a review of an emerging zoonosis". The Journal of General Virology. 90 (Pt 8): 1781–1794. doi:10.1099/vir.0.011437-0. PMID 19420159.
- Goodman JL, Dennis DT, Sonenshine DE (2005). "Tick-Borne Encephalitis". Tick-Borne Diseases of Humans. Washington, DC: ASM Press. p. 151. ISBN 978-1-55581-238-6.
- Kovalev SY, Mukhacheva TA, Kokorev VS, Belyaeva IV (April 2012). "Tick-borne encephalitis virus: reference strain Sofjin and problem of its authenticity". Virus Genes. 44 (2): 217–24. doi:10.1007/s11262-011-0690-9. PMID 22095094.
- Mansfield, K. L.; Johnson, N.; Phipps, L. P.; Stephenson, J. R.; Fooks, A. R.; Solomon, T. (2009-08-01). "Tick-borne encephalitis virus – a review of an emerging zoonosis". Journal of General Virology. 90 (8): 1781–1794. doi:10.1099/vir.0.011437-0. ISSN 0022-1317. PMID 19420159.
- Wilder-Smith, Annelies; Ooi, Eng-Eong; Horstick, Olaf; Wills, Bridget (January 2019). "Dengue". The Lancet. 393 (10169): 350–363. doi:10.1016/s0140-6736(18)32560-1. ISSN 0140-6736. PMID 30696575.
- Ruzek, Daniel; Avšič Županc, Tatjana; Borde, Johannes; Chrdle, Ales; Eyer, Ludek; Karganova, Galina; Kholodilov, Ivan; Knap, Nataša; Kozlovskaya, Liubov; Matveev, Andrey; Miller, Andrew D. (2019-04-01). "Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines". Antiviral Research. 164: 23–51. doi:10.1016/j.antiviral.2019.01.014. ISSN 0166-3542. PMID 30710567.
- Labuda, M.; Jones, L. D.; Nuttall, P. A.; Kaufman, W. R. (1994). "Adaptations of arboviruses to ticks". ERA. doi:10.7939/r3m03xx77. Retrieved 2020-04-17.
- Danielová, Vlasta; Holubová, Jaroslava; Pejcoch, Milan; Daniel, Milan (2002). "Potential significance of transovarial transmission in the circulation of tick-borne encephalitis virus". Folia Parasitologica. 49 (4): 323–325. doi:10.14411/fp.2002.060. ISSN 0015-5683. PMID 12641208.
- Pulkkinen, Lauri; Butcher, Sarah; Anastasina, Maria (2018-06-28). "Tick-Borne Encephalitis Virus: A Structural View". Viruses. 10 (7): 350. doi:10.3390/v10070350. ISSN 1999-4915. PMC 6071267. PMID 29958443.
- Velay, Aurélie; Paz, Magali; Cesbron, Marlène; Gantner, Pierre; Solis, Morgane; Soulier, Eric; Argemi, Xavier; Martinot, Martin; Hansmann, Yves; Fafi-Kremer, Samira (2019-07-04). "Tick-borne encephalitis virus: molecular determinants of neuropathogenesis of an emerging pathogen". Critical Reviews in Microbiology. 45 (4): 472–493. doi:10.1080/1040841X.2019.1629872. ISSN 1040-841X. PMID 31267816.
- Blom, Kim; Cuapio, Angelica; Sandberg, J. Tyler; Varnaite, Renata; Michaëlsson, Jakob; Björkström, Niklas K.; Sandberg, Johan K.; Klingström, Jonas; Lindquist, Lars; Gredmark Russ, Sara; Ljunggren, Hans-Gustaf (2018). "Cell-Mediated Immune Responses and Immunopathogenesis of Human Tick-Borne Encephalitis Virus-Infection". Frontiers in Immunology. 9: 2174. doi:10.3389/fimmu.2018.02174. ISSN 1664-3224. PMC 6168641. PMID 30319632.
- Subbotina EL, Loktev VB (2012). "[Molecular evolution of the tick-borne encephalitis and Powassan viruses]". Molekuliarnaia Biologiia. 46 (1): 82–92. doi:10.1134/S0026893311060148. PMID 22642104.
- Uzcátegui NY, Sironen T, Golovljova I, Jääskeläinen AE, Välimaa H, Lundkvist Å, et al. (April 2012). "Rate of evolution and molecular epidemiology of tick-borne encephalitis virus in Europe, including two isolations from the same focus 44 years apart". The Journal of General Virology. 93 (Pt 4): 786–796. doi:10.1099/vir.0.035766-0. PMID 22205716.
- Suzuki Y (June 2007). "Multiple transmissions of tick-borne encephalitis virus between Japan and Russia". Genes & Genetic Systems. 82 (3): 187–95. doi:10.1266/ggs.82.187. PMID 17660689.
- Takashima I, Hayasaka D, Goto A, Kariwa H, Mizutani T (February 2001). "Epidemiology of tick-borne encephalitis (TBE) and phylogenetic analysis of TBE viruses in Japan and Far Eastern Russia". Japanese Journal of Infectious Diseases. 54 (1): 1–11. PMID 11326122.
- Vene S, Golovljova I, Mavtchoutko V, Forsgren M, Kalnina V, Plyusnin A (December 2001). "Characterization of tick-borne encephalitis virus from Latvia: evidence for co-circulation of three distinct subtypes". Journal of Medical Virology. 65 (4): 730–5. doi:10.1002/jmv.2097. PMID 11745938.
- Golovljova I, Vene S, Sjölander KB, Vasilenko V, Plyusnin A, Lundkvist A (December 2004). "Characterization of tick-borne encephalitis virus from Estonia". Journal of Medical Virology. 74 (4): 580–8. doi:10.1002/jmv.20224. PMID 15484275.
- Mickiené A, Vene S, Golovljova I, Laiskonis A, Lindquist L, Plyusnin A, Lundkvist A (December 2001). "Tick-borne encephalitis virus in Lithuania". European Journal of Clinical Microbiology & Infectious Diseases. 20 (12): 886–8. doi:10.1007/s10096-001-0637-5. PMID 11837641.
- Heinz, Franz-Xaver; Stiasny, Karin (2019). "Chapter 2b: The molecular and antigenic structure of TBEV". Tick-borne Encephalitis - the Book. doi:10.33442/978-981-14-0914-1_2b. ISSN 2661-3980.
- Zlobin, Vladimir I.; Pogodina, Vanda V.; Kahl, Olaf (2017-10-01). "A brief history of the discovery of tick-borne encephalitis virus in the late 1930s (based on reminiscences of members of the expeditions, their colleagues, and relatives)". Ticks and Tick-borne Diseases. 8 (6): 813–820. doi:10.1016/j.ttbdis.2017.05.001. ISSN 1877-959X. PMID 28526419.
- Uspensky, Igor (May 2018). "Several words in addition to "A brief history of the discovery of tick-borne encephalitis virus in the late 1930s" by V.I. Zlobin, V.V. Pogodina and O. Kahl (TTBDIS, 2017, 8, 813–820)". Ticks and Tick-borne Diseases. 9 (4): 834–835. doi:10.1016/j.ttbdis.2018.03.007. PMID 29559213.
External links
- Encephalitis+Viruses,+Tick-Borne at the US National Library of Medicine Medical Subject Headings (MeSH)
- Tick-borne encephalitis at World Health Organization