Caecilian
Caecilians (/sɪˈsɪliən/; New Latin for "blind ones") are a group of limbless, vermiform or serpentine amphibians. They mostly live hidden in the ground and in stream substrates, making them the least familiar order of amphibians. All modern caecilians and their closest fossil relatives are grouped as a clade, Apoda, within the larger group Gymnophiona, which also includes more primitive extinct caecilian-like amphibians.[1] Caecilians are mostly distributed in the tropics of South and Central America, Africa, and southern Asia. Their diet consists of small subterranean creatures such as earthworms.
| Caecilian | |
|---|---|
.jpg) | |
| Oscaecilia ochrocephala | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Amphibia |
| Order: | Gymnophiona |
| Clade: | Apoda Oppel, 1811 |
| Families | |
|
Caeciliidae | |
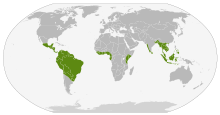 | |
| Current distribution of caecilians (in green) | |
Description
Caecilians completely lack limbs, making the smaller species resemble worms, while the larger species, with lengths up to 1.5 m (5 ft), resemble snakes. Their tails are short or absent, and their cloacae are near the ends of their bodies.[2][3][4]
Their skin is smooth and usually dark, but some species have colourful skins. Inside the skin are calcite scales. Because of these scales, the caecilians were once thought to be related to the fossil Stegocephalia, but they are now believed to be a secondary development, and the two groups are most likely unrelated.[4] The skin also has numerous ring-shaped folds, or annuli, that partially encircle the body, giving them a segmented appearance. Like some other living amphibians, the skin contains glands that secrete a toxin to deter predators.[5] The skin secretions of Siphonops paulensis have been shown to have hemolytic properties.[6]

Caecilians' vision is limited to dark-light perception,[7] and their anatomy is highly adapted for a burrowing lifestyle. They have a strong skull, with a pointed snout used to force their way through soil or mud.[4] In most species, the bones in the skull are reduced in number and fused together, and the mouth is recessed under the head. Their muscles are adapted to pushing their way through the ground, with the skeleton and deep muscles acting as a piston inside the skin and outer muscles. This allows the animal to anchor its hind end in position, and force the head forwards, and then pull the rest of the body up to reach it in waves. In water or very loose mud, caecilians instead swim in an eel-like fashion.[5] Caecilians in the family Typhlonectidae are aquatic, and the largest of their kind. The representatives of this family have a fleshy fin running along the rear section of their bodies, which enhances propulsion in water.[8]
All but the most primitive caecilians have two sets of muscles for closing the jaw, compared with the single pair found in other creatures. These are more highly developed in the most efficient burrowers among the caecilians, and appear to help keep the skull and jaw rigid.[5]
All caecilians possess a pair of tentacles located between their eyes and nostrils. These are probably used for a second olfactory capability, in addition to the normal sense of smell based in the nose.[5]
The ringed caecilian (Siphonops annulatus) has dental glands that may be homologous to the venom glands of some snakes and lizards. The function of these glands is unknown.[9]
The middle ear consists of only the stapes and the oval window, which transfer vibration to the inner ear through a reentrant fluid circuit as seen in some reptiles. The species within the Scolecomorphidae lack both stapes and an oval window, making them the only known amphibians missing all the components of a middle ear apparatus.[10]
Except for one lungless species, Atretochoana eiselti,[11] all caecilians have lungs, but also use their skin or mouths for oxygen absorption. Often, the left lung is much smaller than the right one, an adaptation to body shape that is also found in snakes.
Distribution
Caecilians are found in wet, tropical regions of Southeast Asia, India, Bangladesh, Nepal[12] and Sri Lanka, parts of East and West Africa, the Seychelles Islands in the Indian Ocean, Central America, and in northern and eastern South America. In Africa, caecilians are found from Guinea-Bissau (Geotrypetes) to southern Malawi (Scolecomorphus), with an unconfirmed record from eastern Zimbabwe. They have not been recorded from the extensive areas of tropical forest in central Africa. In South America, they extend through subtropical eastern Brazil well into temperate northern Argentina. They can be seen as far south as Buenos Aires, when they are carried by the flood waters of the Paraná River coming from farther north. Their American range extends north to southern Mexico. The northernmost distribution is of the species Ichthyophis sikkimensis of northern India. Ichthyophis is also found in South China and North Vietnam. In Southeast Asia, they are found as far east as Java, Borneo, and the southern Philippines, but they have not crossed Wallace's line and are not present in Australia or nearby islands. There are no known caecilians in Madagascar, but their presence in the Seychelles and India has posited speculation on the presence of undiscovered extinct or extant caecilians there.[13]
Taxonomy
The name caecilian derives from the Latin word caecus, meaning "blind", referring to the small or sometimes nonexistent eyes. The name dates back to the taxonomic name of the first species described by Carl Linnaeus, which he named Caecilia tentaculata.[4]
There has historically been disagreement over the use of the two primary scientific names for caecilians, Apoda and Gymnophiona. Some specialists prefer to use the name Gymnophiona to refer to the "crown group", that is, the group containing all modern caecilians and extinct members of these modern lineages. They sometimes use the name Apoda to refer to the total group, that is, all caecilians and caecilian-like amphibians that are more closely related to modern groups than to frogs or salamanders. However, many scientists have advocated for the reverse arrangement, where Apoda is used as the name for modern caecilian groups. Some have argued that this use makes more sense, because the name "Apoda" means "without feet", and this is a feature associated mainly with modern species (some stem-group caecilian-like amphibians, such as Eocaecilia, had legs).[14]
The most recent classification of caecilians, by Wilkinson et al. (2011), divided the caecilians into 9 families containing nearly 200 species.[15] Since then, a tenth caecilian family has been discovered, Chikilidae.[16][17] This classification is based on a thorough definition of monophyly based on morphological and molecular evidence,[18][19][20][21] and it solves the longstanding problems of paraphyly of the Caeciliidae in previous classifications without an exclusive reliance upon synonymy.[15][22] It contains 256 species in 56 genera.
- Rhinatrematidae – 2 genera, 13 species; South America
- Ichthyophiidae – 2 genera, 57 species; South and Southeast Asia
- Scolecomorphidae – 2 genera, 6 species; Africa
- Herpelidae – 2 genera, 10 species; Africa
- Chikilidae – 1 genus, 4 species; India
- Caeciliidae – 2 genera, 43 species; South and Central America
- Typhlonectidae – 5 genera, 14 species; South America
- Indotyphlidae – 7 genera, 24 species; Seychelles, India, Africa
- Siphonopidae – 5 genera, 28 species; South America
- Dermophiidae – 4 genera, 14 species; Africa, Central and South America
The most recent phylogeny of caecilians is based on molecular mitogenomic evidence examined by San Mauro et al. (2014).[23][24]
| Gymnophiona |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Evolution

Little is known of the evolutionary history of the caecilians, which have left a very sparse fossil record. The first fossil, a vertebra dated to the Paleocene, was not discovered until 1972.[25] Other vertebrae, which have characteristic features unique to modern species, were later found in Paleocene and Late Cretaceous (Cenomanian) sediments.[1]
The earliest fossil attributed to a stem-caecilian (a species closer to caecilians than to frogs or salamanders but not a member of the extant lineage) comes from the Jurassic period. This primitive genus, Eocaecilia, had small limbs and well-developed eyes.[26] In their 2008 description of the fossil batrachian Gerobatrachus,[27] Anderson and co-authors suggested that caecilians arose from the Lepospondyl group of ancestral tetrapods, and may be more closely related to amniotes than to frogs and salamanders, which arose from Temnospondyl ancestors. Numerous groups of lepospondyls evolved reduced limbs, elongated bodies, and burrowing behaviors, and morphological studies on Permian and Carboniferous lepospondyls have placed the early caecilian (Eocaecilia) among these groups.[28] Divergent origins of caecilians and other extant amphibians may help explain the slight discrepancy between fossil dates for the origins of modern amphibia, which suggest Permian origins, and the earlier dates, in the Carboniferous, predicted by some molecular clock studies of DNA sequences. Most morphological and molecular studies of extant amphibians, however, support monophyly for caecilians, frogs, and salamanders, and the most recent molecular study based on multi-locus data suggest a Late Carboniferous–Early Permian origin of extant amphibians.[29] The proposed molecular clock for the origin of lissamphibians has been further strengthened by the discovery of Chinlestegophis from the Late Triassic Chinle Formation of Colorado.[30]
Behavior
Reproduction
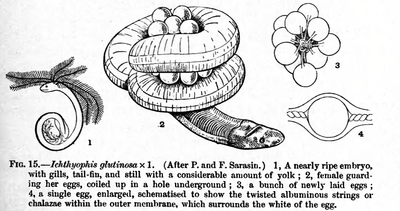
Caecilians are the only order of amphibians to use internal insemination exclusively (although most salamanders have internal fertilization and the tailed frog in the US uses a tail-like appendage for internal insemination in its fast-flowing water environment).[5] The male caecilians have a long tube-like intromittent organ, the phallodeum,[31] which is inserted into the cloaca of the female for two to three hours. About 25% of the species are oviparous (egg-laying); the eggs are guarded by the female. For some species, the young caecilians are already metamorphosed when they hatch; others hatch as larvae. The larvae are not fully aquatic, but spend the daytime in the soil near the water.[5]
About 75% of caecilians are viviparous, meaning they give birth to already-developed offspring. The foetus is fed inside the female with cells lining the oviduct, which they eat with special scraping teeth.
The egg-laying species Boulengerula taitana feeds its young by developing an outer layer of skin, high in fat and other nutrients, which the young peel off with modified teeth. This allows them to grow by up to 10 times their own weight in a week. The skin is consumed every three days, the time it takes for a new layer to grow, and the young have only been observed to eat it at night. It was formerly thought that the juveniles subsisted only on a liquid secretion from their mothers.[32][33]
Some larvae, such as those of Typhlonectes, are born with enormous external gills which are shed almost immediately.
Diet
The diets of caecilians are not well known. Mature caecilians seem to feed mostly on insects and other invertebrates found in the habitat of the respective species. The stomach contents of 14 specimens of Boulengerula taitana consisted of mostly unidentifiable organic material and plant remains. Where identifiable remains were most abundant, they were found to be termite heads.[34] While the undefinable organic material may show the caecilians eat detritus, the remains may be from earthworms. Caecilians in captivity can be easily fed with earthworms, and worms are also common in the habitat of many caecilian species.
See also
- Caecilians of the Western Ghats
- Minhocão – a cryptid that resembles caecilians
References
Specific references:
- Evans, Susan E.; Sigogneau-Russell, Denise (2001). "A stem-group caecilian (Lissamphibia: Gymnophiona) from the Lower Cretaceous of North Africa". Palaeontology. 44 (2): 259–273. doi:10.1111/1475-4983.00179.
- Goin, C. J.; Goin, O.B.; Zug, G.W. (1978). "Order Gymnophiona". Introduction to Herpetology (3rd ed.). San Francisco: W.H. Freeman and Company. p. 201. ISBN 978-0-7167-0020-3.
- Himstedt, Werner (1996). Die Blindwühlen (in German). Magdeburg: Westarp Wissenschaften. ISBN 978-3-89432-434-6.
- Chisholm, Hugh, ed. (1911). . Encyclopædia Britannica (11th ed.). Cambridge University Press.
- Nussbaum, Ronald A. (1998). Cogger, H.G.; Zweifel, R.G. (eds.). Encyclopedia of Reptiles and Amphibians. San Diego: Academic Press. pp. 52–59. ISBN 978-0-12-178560-4.
- Elisabeth N. Ferroni Schwartz; Carlos A. Schwartz; Antonio Sebben (1998). "Occurrence of hemolytic activity in the skin secretion of the caecilian Siphonops paulensis". Natural Toxins. 6 (5): 179–182. doi:10.1002/(SICI)1522-7189(199809/10)6:5<179::AID-NT20>3.0.CO;2-M. PMID 10398514.
- Mohun, S. M.; Davies, W. L.; Bowmaker, J. K.; Pisani, D.; Himstedt, W.; Gower, D. J.; Hunt, D. M.; Wilkinson, M. (2010). "Identification and characterization of visual pigments in caecilians (Amphibia: Gymnophiona), an order of limbless vertebrates with rudimentary eyes". The Journal of Experimental Biology. 213 (20): 3586–3592. doi:10.1242/jeb.045914. PMID 20889838.
- Piper, Ross (2007). Extraordinary Animals: An Encyclopedia of Curious and Unusual Animals. Greenwood Press.
- Mailho-Fontana, Pedro Luiz; Antoniazzi, Marta Maria; Alexandre, Cesar; Pimenta, Daniel Carvalho; Sciani, Juliana Mozer; Brodie, Edmund D.; Jared, Carlos (July 2020). "Morphological Evidence for an Oral Venom System in Caecilian Amphibians". iScience: 101234. doi:10.1016/j.isci.2020.101234. ISSN 2589-0042.
- Hearing and Sound Communication in Amphibians
- "Atretochoana eiselti". Natural History Museum. Retrieved 22 February 2012.
- Rathor, Hariharsingh (5 May 2016). "Kutnjema Mareka Jiv Sarpa Hoina". Katipur. Kantipur News. Retrieved 5 May 2016.
- James D. Gardner, Jean-Claude Rage, The fossil record of lissamphibians from Africa, Madagascar, and the Arabian Plate, Palaeobiodiversity and Palaeoenvironments 96(1):1-52 · March 2016
- Marjanovic, D.; Laurin, M. (2007). "Fossils, molecules, divergence times, and the origin of lissamphibians". Syst. Biol. 56 (3): 369–388. doi:10.1080/10635150701397635. PMID 17520502.
- Wilkinson, M.; San Mauro, D.; Sherratt, E.; Gower, D.J. (2011). "A nine-family classification of caecilians (Amphibia: Gymnophiona)" (PDF). Zootaxa. 2874: 41–64. doi:10.11646/zootaxa.2874.1.3.
- Kamei, R.G.; San Mauro, D.; Gower, D. J.; Van Bocxlaer, I.; Sherratt, E.; Thomas, A.; Babu, S.; Bossuyt, F.; Wilkinson, M.; Biju, S. D. (2012). "Discovery of a new family of amphibians from Northeast India with ancient links to Africa". Proc. R. Soc. B. 279 (1737): 2396–401. doi:10.1098/rspb.2012.0150. PMC 3350690. PMID 22357266.
- "New amphibian family found in India". CBC News. Associated Press. 21 February 2012.
- San Mauro, D.; Gower, D. J.; Oommen, O. V.; Wilkinson, M.; Zardoya, R. (2004). "Phylogeny of caecilian amphibians (Gymnophiona) based on complete mitochondrial genomes and nuclear RAG1". Molecular Phylogenetics and Evolution. 33 (2): 413–427. doi:10.1016/j.ympev.2004.05.014. PMID 15336675.
- San Mauro, D.; Gower, D. J.; Massingham, T.; Wilkinson, M.; Zardoya, R.; Cotton, J. A. (2009). "Experimental design in caecilian systematics: phylogenetic information of mitochondrial genomes and nuclear rag1". Systematic Biology. 58 (4): 425–438. CiteSeerX 10.1.1.577.2856. doi:10.1093/sysbio/syp043. PMID 20525595.
- Zhang, P.; Wake, M. H. (2009). "A mitogenomic perspective on the phylogeny and biogeography of living caecilians (Amphibia: Gymnophiona)". Molecular Phylogenetics and Evolution. 53 (2): 479–491. doi:10.1016/j.ympev.2009.06.018. PMID 19577653.
- San Mauro, D.; Gower, D. J.; Cotton, J. A.; Zardoya, R.; Wilkinson, M.; Massingham, T. (2012). "Experimental design in phylogenetics: testing predictions from expected information". Systematic Biology. 61 (4): 661–674. doi:10.1093/sysbio/sys028. PMID 22328568.
- Frost, Darrel R.; Grant, Taran; Faivovich, Julián; Bain, Raoul H.; Haas, Alexander; Haddad, Célio F.B.; De Sá, Rafael O.; Channing, Alan; Wilkinson, Mark; Donnellan, Stephen C.; Raxworthy, Christopher J.; Campbell, Jonathan A.; Blotto, Boris L.; Moler, Paul; Drewes, Robert C.; Nussbaum, Ronald A.; Lynch, John D.; Green, David M.; Wheeler, Ward C. (2006). "The Amphibian Tree of Life". Bulletin of the American Museum of Natural History. 297: 1–370, appendices. doi:10.1206/0003-0090(2006)297[0001:TATOL]2.0.CO;2. ISSN 0003-0090.
- Pyron, R.A.; Wiens, J.J. (2011). "A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians". Molecular Phylogenetics and Evolution. 61 (2): 543–583. doi:10.1016/j.ympev.2011.06.012. PMID 21723399.
- San Mauro, D.; Gower, D. J.; Müller, H.; Loader, S. P.; Zardoya, R.; Nussbaum, R. A.; Wilkinson, M. (2014). "Life-history evolution and mitogenomic phylogeny of caecilian amphibians". Molecular Phylogenetics and Evolution. 73: 177–89. doi:10.1016/j.ympev.2014.01.009. hdl:10261/123960. PMID 24480323.
- Estes, Richard; Wake, Marvalee H. (22 September 1972). "The First Fossil Record of Caecilian Amphibians". Nature. 239 (5369): 228–231. Bibcode:1972Natur.239..228E. doi:10.1038/239228b0.
- Jenkins, Parish A.; Walsh, Denis M. (16 September 1993). "An Early Jurassic caecilian with limbs". Nature. 365 (6443): 246–250. Bibcode:1993Natur.365..246J. doi:10.1038/365246a0.
- Anderson, Jason S.; Reisz, Robert R.; Scott, Diane; Fröbisch, Nadia B.; Sumida, Stuart S. (2008). "A stem batrachian from the Early Permian of Texas and the origin of frogs and salamanders". Nature. 453 (7194): 515–8. Bibcode:2008Natur.453..515A. doi:10.1038/nature06865. PMID 18497824.
- Huttenlocker, A. K.; Pardo, J. D.; Small, B. J.; Anderson, J. S. (2013). "Cranial morphology of recumbirostrans (Lepospondyli) from the Permian of Kansas and Nebraska, and early morphological evolution inferred by micro-computed tomography". Journal of Vertebrate Paleontology. 33 (3): 540–552. doi:10.1080/02724634.2013.728998.
- San Mauro, D. (2010). "A multilocus timescale for the origin of extant amphibians". Molecular Phylogenetics and Evolution. 56 (2): 554–561. doi:10.1016/j.ympev.2010.04.019. PMID 20399871.
- Jason D. Pardo; Bryan J. Small; Adam K. Huttenlocker (2017). "Stem caecilian from the Triassic of Colorado sheds light on the origins of Lissamphibia". Proceedings of the National Academy of Sciences of the United States of America. in press. doi:10.1073/pnas.1706752114.
- Barrie G M Jamieson (5 January 2006). Reproductive Biology and Phylogeny of Gymnophiona: Caecilians. CRC Press. ISBN 978-1-4822-8014-2.
- Kupfer, Alex; Muller, Hendrik; Antoniazzi, Marta M.; Jared, Carlos; Greven, Hartmut; Nussbaum, Ronald A.; Wilkinson, Mark (2006). "Parental investment by skin feeding in a caecilian amphibian" (PDF). Nature. 440 (7086): 926–929. Bibcode:2006Natur.440..926K. doi:10.1038/nature04403. hdl:2027.42/62957. PMID 16612382.
- Vince, Gaia (12 April 2006). "'Yummy mummy' worms feed their skin to offspring". New Scientist.
- Hebrard, J. J.; Maloiy, G. M. O.; Al-Liangana, D. M. I. (1992). "Notes on the habitat and diet of Afrocaecilia taitana". Journal of Herpetology. 26 (4): 513–515. doi:10.2307/1565136. JSTOR 1565136.
External links
| Wikispecies has information related to Gymnophiona |
| Wikimedia Commons has media related to Gymnophiona. |