Teleost leptins
Teleost leptins are a family of peptide hormones found in fish (teleostei) that are orthologs of the mammalian hormone leptin. The teleost and mammalian leptins appear to have similar functions, namely, regulation of energy intake and expenditure.
| Leptin | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
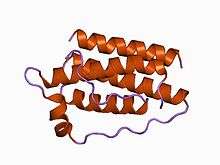 Structure of the obese protein leptin-E100.[1] | |||||||||||
| Identifiers | |||||||||||
| Symbol | Leptin | ||||||||||
| Pfam | PF02024 | ||||||||||
| Pfam clan | CL0053 | ||||||||||
| InterPro | IPR000065 | ||||||||||
| SCOPe | 1ax8 / SUPFAM | ||||||||||
| |||||||||||
The leptin (LEP) hormone was long thought to be specific to mammals, but in recent years the gene (lep) has been found in amphibia such as the tiger salamander (Ambystoma tigrinum),[2] and the African clawed frog (Xenopus laevi).[3] The discovery of lep in puffer fish (Takifugu rubripes)[4] demonstrates the ancient ancestry of this hormone.
Examples
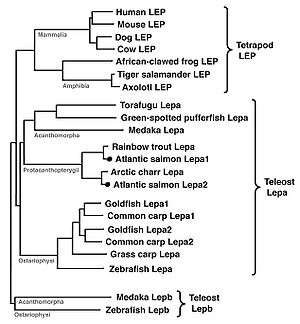
There are two closely related lep paralogues in Atlantic salmon (Salmo salar).[5] A single lep gene has been documented for green-spotted pufferfish (Tetraodon nigroviridis),[4] rainbow trout (Oncorhynchus mykiss),[6] Arctic charr (Salvelinus alpinus),[7] silver carp (Hypophthalmichthys molitrix), and grass carp (Ctenopharyngodon idellus).[8] In other species there are reports of two closely related lep paralogues, including common carp (Cyprinus carpio)[9] and Atlantic salmon.[5] More distantly related lep genes have been found in medaka (Oryzias latipes)[10] and zebrafish (Danio rerio).[11] At least 2 leptin genes (lepa and lepb) exist in the crown-clade (Fig. 1).[4][9][10][11] Early findings have shown that lepa and lepb share low interspecies aa identity, and are argued to have arisen through whole genome duplication,[4] which occurred early in the teleost lineage.[12] The duplicity of genes has been described for Atlantic salmon, Japanese medaka, common carp and zebrafish.[5][9][10][11] Both lep paralogues[5] cluster with lepa, and therefore suggest that at least one or more form (lepb) may exist in this species, since it is tetraploid.[13] However, previous attempts using genomic synteny have only found the putative genomic duplicates in medaka and zebrafish paralogue.[10][11] Currently it remains unclear, whether lepb exists in other teleosts due to the degenerative nature of this paralogue.
Comparison with mammalian leptin
The large differences among endothermic (warm-blooded) mammalian and ectothermic (cold-blooded) teleost leptins raised the question of whether the energy homeostatic functions of the teleost leptins are conserved. Initial phylogenetic analysis has revealed that amino acid conservation with other vertebrate Lep orthologues is low, with only 13.2% sequence identity between torafugu and human LEP.[4] Subsequent investigations have confirmed the low amino acid identity of teleost leps compared to mammalian LEP.[4][9][14][15]
Structure
The three-dimensional homology modeling predicts strong conservation of the tertiary structure between Atlantic salmon and other teleost Leps compared to their mammalian orthologues (Fig. 2).[4][5][6][10][11]
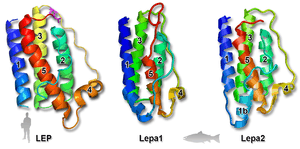
Both lepa1 and lepa2 have two characteristic cysteine residues which predict the formation of a disulfide bond in Lep, which is a pre-equisite for this 3D configuration and bioactivity of human LEP.[18][19] The models suggest that the bonding of lepa2 might be different from lepa1. There are several differences between the 3D structures of lepa1 and lepa2; e.g. α-helix 5 is considerable shorter in lepa1 than lepa2. Furthermore, α-helix 1 for lepa2 appears to be split by a short-disordered region, and may therefore have a poorer affinity. However, considering that it is a predicted model based upon the structure mask of human LEP, the significance of these putative conformational adjustments remains to be tested.
The importance of the conserved tertiary structure of Lep is most likely explained by requirements for specific LepR-binding affinity and is constrained by the structure of the receptor-binding pocket.[3] This might also explain some of the results from studies on teleost using heterologous mammalian Lep. E.g. treatment with the mammalian hormone caused an anorexic effect in goldfish (Carassius auratus)[20][21] and green sunfish (Lepomis cyanellus),[22] but not in Coho salmon (Oncorhynchus kisutch),[23] channel catfish (Ictalurus punctatus) [24] and green sunfish.[25] These contradicting results have been explained by the relatively large differences in amino acid sequences observed between mammals and fish.[3][4][9]
Rønnestad and colleagues[5] recently detected five isoforms of the leptin receptor (lepr) that have differences in 3'-end of the mRNA sequence. Of these, only the longest form conserved all functionally important domains (such as three fibronectin type III domains, the Ig C2-like domain, a pair of WSXWS motifs, two JAK2-binding motif boxes, and a STAT-binding domain),[5] while the other four forms have only the intra-cellular region. The long form of mammalian LepR has a function for full signal transduction through the JAK/STAT pathways, whereas the shorter forms exhibit partial or no signaling capabilities.[26][27] The biological importance of long form LepR via the JAK/STAT pathway in maintaining body weight and energy homeostasis has been demonstrated.[28] Previous studies in teleosts have only identified a single lepr.[10][14][29] Rønnestad et al.,[5] is the first to report that plural LepR transcripts in any ectotherm species. When looking at the available motif for lepr, the model suggests that it would bind easily to lepa1 and not lepa2 (Fig. 2). Furthermore, the relatively ubiquitous expression of lepr in salmon tissues supports diverse roles of lep in teleosts.[5]
Tissue distribution

The study on torafugu[4] indicated that lep is mainly expressed in the liver in contrasts to the adipose secretion in mammals.[30][31] However recent studies have shown that lep is expressed in several peripheral tissues, including intestine, kidney, ovary, muscle and adipose tissue.[5][6][10] The multiplicity of lep genes and their low conservation in Teleostei.[9][10][11] suggest that their physiological roles may be more divergent than reported for mammals.
The tissue expression pattern for the Atlantic salmon lep paralogues differs substantially (Fig.3)[5] and hence indicates a possible difference in function. With the exception of the results presented here, and those for zebrafish and Japanese medaka.[5][10][11] Few studies have investigated the broad tissue distribution of lep in teleost fishes. The more distantly related lep genes (lepa and lepb) showed distinct differences in tissue distribution, as shown in e.g. medaka, where lepa is being expressed in liver and muscle, while lepb is more highly expressed in the brain and eye. However, these differences are also observed for more closely related lep paralogues, such as lepa1 in Atlantic salmon, being more highly expressed in brain, liver and white muscle, while lepa2 is mainly expressed in the stomach and midgut. (Fig. 3).
Effects of nutritional status
The observations that long-term feed restriction does not significantly affect lep expression in Atlantic salmon[5] has also been noted in other teleosts. However it is likely that prolonged feed restriction can influence several endocrine parameters to adapt to the nutritional condition. For example, in common carp, a rapid response in ob gene expression in hepatic tissue of common carp shortly after feeding, but no changes in expression in response to different long-term feeding regime was observed.[9] These authors suggested that this effect could be explained by the fact that starved fish do not lose weight as rapidly as mammals, a consequence of being ectothermic and possessing a much lower standard metabolic rate, and therefore can withstand longer periods of starvation. A similar study on grass carp showed that chronic injection of species-specific Lep did not affect long-term food intake and body weight, while acute injection decreased food intake.[8] Conversely, Murashita et al., (unpublished results) observed increased proopiomelanocortin a1(pomca1) levels following chronic injection of Lep in Atlantic salmon, which suggests that chronic exposure to elevated Lep levels can decrease food intake through the Pomc pathway in this species. Recent studies in Atlantic salmon did not observe any difference of feed restriction in circulating plasma levels,[17] which contrasts recent results in rainbow trout[32] and suggest that the relation between circulating lep levels and energy status differs from that in mammals. However, salmon RIA appears to allow interspecies assessment of plasma lep levels.[32] This only confirms that more comprehensive studies are needed for conclusive data interpretation. Studies on rainbow trout also implicated Lep as an anorectic hormone as in mammals. Injection of rainbow trout with recombinant trout leptin (rt-leptin) resulted in a significantly reduced appetite over two days that coincided with a decrease in hypothalamic mRNA expression of neuropeptide Y (npy) and increase of pomc mRNAs, respectively.[6] Whether these observations are due to species-specific differences in long-term leptin regulation of appetite or growth is not known, however, consensus data indicate that the effects of Lep on appetite regulation may be short-term in teleosts.
Short-term feed restriction
Recent studies on short-term effects of a meal or the absence of a meal[17] has revealed that lepa1 expression specifically peaks in the peripheral tissues after 6 – 9 hr in the unfed fish. This suggests that the transcript specific response could be associated with the absence of food. Conversely, since the unfed fish had not received food for 33 hr (24 + 9 hr), the peaks could represent an unrelated effect. Each lepa1 peak occurred during a phase of falling plasma Lep, and since this occurred in both fed and unfed fish, the temporal upregulation of lepa1 does not in fact appear to be specifically related to the absence of food.
The earliest peak of lepa1 occurred in the white muscle, which represents an important lipid reservoir in Atlantic salmon.[33] Unlike pufferfish, which utilizes the liver as a major lipid repository,[4] Atlantic salmon shows that despite a high visceral lipid content, hepatocytes contain few lipid droplets compared to other fish species,[34] yet are an important site for leptin expression.[4][5][6][9][11][17] Moen and colleagues[17] reported that both lepa1 and lepa2 peaked at 9 hr in the liver of unfed fish. By contrast, however, studies in common carp demonstrated a peak in leptin-I(lepa1) and leptin–II (lepa2) in liver at 3 and 6 hr post feeding respectively.[9] The earlier expression response of leptins in common carp likely reflects the higher temperature under which the experiments were conducted, but contrasts the findings of upregulation of lepa1 due to the absence of food.[17] Similarly, in mice, a postprandial increase in hepatic leptin expression has also been reported.[35] However, in grass carp, intraperitoneal injection of recombinant leptin only alters the appetite on the first day, and does not influence food intake during the ensuing 12 days.[15] At present, the data for Atlantic salmon are therefore quite different and suggest that leptin expression in this species may have a complex lipostatic function.
References
- Zhang F, Basinski MB, Beals JM, et al. (May 1997). "Crystal structure of the obese protein leptin-E100". Nature. 387 (6629): 206–9. doi:10.1038/387206a0. PMID 9144295.
- Boswell T, Dunn IC, Wilson PW, Joseph N, Burt DW, Sharp PJ (April 2006). "Identification of a non-mammalian leptin-like gene: characterization and expression in the tiger salamander (Ambystoma tigrinum)" (PDF). Gen. Comp. Endocrinol. 146 (2): 157–66. doi:10.1016/j.ygcen.2005.08.001. hdl:20.500.11820/4d690bb6-fa7b-4d57-ae9d-98f3dfa1aebc. PMID 16480984.
- Crespi EJ, Denver RJ (June 2006). "Leptin (ob gene) of the South African clawed frog Xenopus laevis". Proc. Natl. Acad. Sci. U.S.A. 103 (26): 10092–7. doi:10.1073/pnas.0507519103. PMC 1502511. PMID 16782821.
- Kurokawa T, Uji S, Suzuki T (May 2005). "Identification of cDNA coding for a homologue to mammalian leptin from pufferfish, Takifugu rubripes". Peptides. 26 (5): 745–50. doi:10.1016/j.peptides.2004.12.017. PMID 15808904.
- Rønnestad I, Nilsen TO, Murashita K, Angotzi AR, Gamst Moen AG, Stefansson SO, Kling P, Thrandur Björnsson B, Kurokawa T (2010). "Leptin and leptin receptor genes in Atlantic salmon: Cloning, phylogeny, tissue distribution and expression correlated to long-term feeding status". Gen. Comp. Endocrinol. 168 (1): 55–70. doi:10.1016/j.ygcen.2010.04.010. hdl:1956/4036. PMID 20403358.
- Murashita K, Uji S, Yamamoto T, Rønnestad I, Kurokawa T (August 2008). "Production of recombinant leptin and its effects on food intake in rainbow trout (Oncorhynchus mykiss)". Comp. Biochem. Physiol. B, Biochem. Mol. Biol. 150 (4): 377–84. doi:10.1016/j.cbpb.2008.04.007. PMID 18539064.
- Frøiland E, Murashita K, Jørgensen EH, Kurokawa T (January 2010). "Leptin and ghrelin in anadromous Arctic charr: cloning and change in expressions during a seasonal feeding cycle". Gen. Comp. Endocrinol. 165 (1): 136–43. doi:10.1016/j.ygcen.2009.06.010. PMID 19539626.
- Li X, He J, Hu W, Yin Z (June 2009). "The essential role of endogenous ghrelin in growth hormone expression during zebrafish adenohypophysis development". Endocrinology. 150 (6): 2767–74. doi:10.1210/en.2008-1398. PMID 19264876.
- Huising MO, Geven EJ, Kruiswijk CP, Nabuurs SB, Stolte EH, Spanings FA, Verburg-van Kemenade BM, Flik G (December 2006). "Increased leptin expression in common Carp (Cyprinus carpio) after food intake but not after fasting or feeding to satiation". Endocrinology. 147 (12): 5786–97. doi:10.1210/en.2006-0824. PMID 16935838.
- Kurokawa T, Murashita K (April 2009). "Genomic characterization of multiple leptin genes and a leptin receptor gene in the Japanese medaka, Oryzias latipes". Gen. Comp. Endocrinol. 161 (2): 229–37. doi:10.1016/j.ygcen.2009.01.008. PMID 19523397.
- Gorissen M, Bernier NJ, Nabuurs SB, Flik G, Huising MO (June 2009). "Two divergent leptin paralogues in zebrafish (Danio rerio) that originate early in teleostean evolution". J. Endocrinol. 201 (3): 329–39. doi:10.1677/JOE-09-0034. PMID 19293295.
- Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N, Mauceli E, Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A, Nicaud S, Jaffe D, Fisher S, Lutfalla G, Dossat C, Segurens B, Dasilva C, Salanoubat M, Levy M, Boudet N, Castellano S, Anthouard V, Jubin C, Castelli V, Katinka M, Vacherie B, Biémont C, Skalli Z, Cattolico L, Poulain J, De Berardinis V, Cruaud C, Duprat S, Brottier P, Coutanceau JP, Gouzy J, Parra G, Lardier G, Chapple C, McKernan KJ, McEwan P, Bosak S, Kellis M, Volff JN, Guigó R, Zody MC, Mesirov J, Lindblad-Toh K, Birren B, Nusbaum C, Kahn D, Robinson-Rechavi M, Laudet V, Schachter V, Quétier F, Saurin W, Scarpelli C, Wincker P, Lander ES, Weissenbach J, Roest Crollius H (October 2004). "Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype". Nature. 431 (7011): 946–57. doi:10.1038/nature03025. PMID 15496914.
- Allendorf FW, Thorgaard G (1984). "Tetraploidy and the evolution of salmonid fishes". In Turner BJ (ed.). Evolutionary genetics of fishes. New York: Plenum Press. pp. 1–45. ISBN 978-0-306-41520-3.
- Kurokawa T, Murashita K, Suzuki T, Uji S (August 2008). "Genomic characterization and tissue distribution of leptin receptor and leptin receptor overlapping transcript genes in the pufferfish, Takifugu rubripes". Gen. Comp. Endocrinol. 158 (1): 108–14. doi:10.1016/j.ygcen.2008.06.003. PMID 18582469.
- Li GG, Liang XF, Xie Q, Li G, Yu Y, Lai K (March 2010). "Gene structure, recombinant expression and functional characterization of grass carp leptin". Gen. Comp. Endocrinol. 166 (1): 117–27. doi:10.1016/j.ygcen.2009.10.009. PMID 19857495.
- Guex N, Peitsch MC, Schwede T (June 2009). "Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: a historical perspective". Electrophoresis. 30 Suppl 1: S162–73. doi:10.1002/elps.200900140. PMID 19517507.
- Moen AG, Finn RN (2013). "Short-term, but not long-term feed restriction causes differential expression of leptins in Atlantic salmon". Gen. Comp. Endocrinol. 183: 83–8. doi:10.1016/j.ygcen.2012.09.027. PMID 23201187.
- York IA, Rock KL (1996). "Antigen processing and presentation by the class I major histocompatibility complex". Annu. Rev. Immunol. 14: 369–96. doi:10.1146/annurev.immunol.14.1.369. PMID 8717519.
- Zhang Y, Olbort M, Schwarzer K, Nuesslein-Hildesheim B, Nicolson M, Murphy E, Kowalski TJ, Schmidt I, Leibel RL (November 1997). "The leptin receptor mediates apparent autocrine regulation of leptin gene expression". Biochem. Biophys. Res. Commun. 240 (2): 492–5. doi:10.1006/bbrc.1997.7622. PMID 9388507.
- de Pedro N, Martínez-Alvarez R, Delgado MJ (March 2006). "Acute and chronic leptin reduces food intake and body weight in goldfish (Carassius auratus)". J. Endocrinol. 188 (3): 513–20. doi:10.1677/joe.1.06349. PMID 16522731.
- Volkoff H, Eykelbosh AJ, Peter RE (May 2003). "Role of leptin in the control of feeding of goldfish Carassius auratus: interactions with cholecystokinin, neuropeptide Y and orexin A, and modulation by fasting". Brain Res. 972 (1–2): 90–109. doi:10.1016/S0006-8993(03)02507-1. PMID 12711082.
- Johnson RM, Johnson TM, Londraville RL (June 2000). "Evidence for leptin expression in fishes". J. Exp. Zool. 286 (7): 718–24. doi:10.1002/(sici)1097-010x(20000601)286:7<718::aid-jez6>3.0.co;2-i. PMC 3506126. PMID 10797324.
- Baker DM, Larsen DA, Swanson P, Dickhoff WW (April 2000). "Long-term peripheral treatment of immature coho salmon (Oncorhynchus kisutch) with human leptin has no clear physiologic effect". Gen. Comp. Endocrinol. 118 (1): 134–8. doi:10.1006/gcen.1999.7450. PMID 10753575.
- Silverstein JT, Plisetskaya EM (2000). "The effects of NPY and insulin on food intake regulation in fish". American Zoologist. 40 (2): 296–308. doi:10.1093/icb/40.2.296.
- Londraville RL, Duvall CS (October 2002). "Murine leptin injections increase intracellular fatty acid-binding protein in green sunfish (Lepomis cyanellus)". Gen. Comp. Endocrinol. 129 (1): 56–62. doi:10.1016/s0016-6480(02)00510-5. PMID 12409096.
- Baumann H, Morella KK, White DW, Dembski M, Bailon PS, Kim H, Lai CF, Tartaglia LA (August 1996). "The full-length leptin receptor has signaling capabilities of interleukin 6-type cytokine receptors". Proc. Natl. Acad. Sci. U.S.A. 93 (16): 8374–8. doi:10.1073/pnas.93.16.8374. PMC 38678. PMID 8710878.
- Tartaglia LA (March 1997). "The leptin receptor". J. Biol. Chem. 272 (10): 6093–6. doi:10.1074/jbc.272.10.6093. PMID 9102398.
- Bates SH, Stearns WH, Dundon TA, Schubert M, Tso AW, Wang Y, Banks AS, Lavery HJ, Haq AK, Maratos-Flier E, Neel BG, Schwartz MW, Myers MG (February 2003). "STAT3 signalling is required for leptin regulation of energy balance but not reproduction". Nature. 421 (6925): 856–9. doi:10.1038/nature01388. PMID 12594516.
- Liu Q, Chen Y, Copeland D, Ball H, Duff RJ, Rockich B, Londraville RL (April 2010). "Expression of leptin receptor gene in developing and adult zebrafish". Gen. Comp. Endocrinol. 166 (2): 346–55. doi:10.1016/j.ygcen.2009.11.015. PMC 3408649. PMID 19941865.
- Montague CT, Farooqi IS, Whitehead JP, Soos MA, Rau H, Wareham NJ, Sewter CP, Digby JE, Mohammed SN, Hurst JA, Cheetham CH, Earley AR, Barnett AH, Prins JB, O'Rahilly S (June 1997). "Congenital leptin deficiency is associated with severe early-onset obesity in humans". Nature. 387 (6636): 903–8. doi:10.1038/43185. PMID 9202122.
- Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM (December 1994). "Positional cloning of the mouse obese gene and its human homologue". Nature. 372 (6505): 425–32. doi:10.1038/372425a0. PMID 7984236.
- Kling P, Rønnestad I, Stefansson SO, Murashita K, Kurokawa T, Björnsson BT (July 2009). "A homologous salmonid leptin radioimmunoassay indicates elevated plasma leptin levels during fasting of rainbow trout". Gen. Comp. Endocrinol. 162 (3): 307–12. doi:10.1016/j.ygcen.2009.04.003. PMID 19362558.
- Aursand M, Bleivik B, Rainuzzo JR, Jørgensen L, Mohr V (1994). "Lipid distribution and composition of commercially farmed Atlantic salmon (Salmo salar)". J. Sci. Food Agric. 64 (2): 239–248. doi:10.1002/jsfa.2740640214.
- Bruslé J., Gonzàlez i Anadon G (1996). "The structure and function of fish liver". In Datta-Munshi JS, Dutta HM (ed.). Fish Morphology. Boca Raton: CRC. pp. 77–93. ISBN 978-90-5410-289-2.
- Saladin R, De Vos P, Guerre-Millo M, Leturque A, Girard J, Staels B, Auwerx J (October 1995). "Transient increase in obese gene expression after food intake or insulin administration". Nature. 377 (6549): 527–9. doi:10.1038/377527a0. PMID 7566150.
| About fish |
|  | |||
|---|---|---|---|---|---|
| Anatomy and physiology | |||||
| Sensory systems | |||||
| Reproduction | |||||
| Locomotion | |||||
| Other behaviour | |||||
| By habitat | |||||
| Other types | |||||
| Commercial |
| ||||
| Major groups | |||||
| Lists |
| ||||
| |||||