Paraspeckle
In cell biology, a Paraspeckle is an irregularly shaped compartment of the cell, approximately 0.2-1 μm in size,[1] found in the nucleus' interchromatin space.[2] First documented in HeLa cells, where there are generally 10-30 per nucleus,[3] Paraspeckles are now known to also exist in all human primary cells, transformed cell lines and tissue sections.[4] Their name is derived from their distribution in the nucleus; the "para" is short for parallel and the "speckle" refers to the splicing speckles to which they are always in close proximity.[3] Their function is still not fully understood, but they are thought to regulate gene expression by sequestrating proteins or mRNAs with inverted repeats in their 3′ UTRs.[5][6]
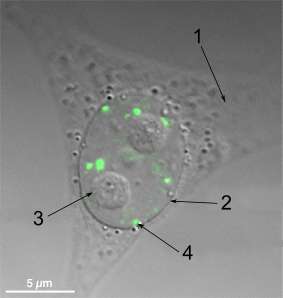
Structure
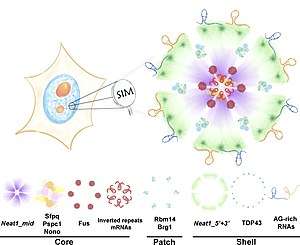
Paraspeckles are organised into core-shell spheroidal structures; seven proteins on a scaffold of lncRNA NEAT1 (the 23kb isoform termed NEAT1_2 or NEAT1v2).[8] In 2016, West et al. proposed the currently accepted model for Paraspeckles. This was based on their current findings using super-resolution microscopy.[7] Their models state that the NEAT1_2 scaffold folds into a V-shaped unit. Many of these units then are assembled into a core-shell spheroid by FUS proteins. Core proteins SFPQ, NONO and PSPC1 tightly associate to the assembled structure. Finally, the shell forms, composed of partially co-localised TDP43 proteins. Due to the integral nature of NEAT1 to paraspeckles assembly, assembly is thought to occur in close proximity to NEAT1 transcription sites.[8]
It has been noted that Paraspeckles have a great deal of commonality both in features and structures with cytoplasmic stress granules, another type of membrane-less organelle. This conclusion arose from the fact that both contain common component proteins,[9] become more abundant with stress, seem to function through sequestering other proteins and both have distinct core or shell regions with predictable localised molecules.[9]
Localization
Paraspeckles are dynamic structures that are altered in response to changes in cellular metabolic activity. They are transcription-dependent.[2] All five of the proposed protein components have RNA recognition motifs (RRMs)[3] and, in the absence of RNA polymerase II transcription, the Paraspeckle disappears and all of its associated proponents form a crescent shaped perinucleolar cap in the nucleolus. This phenomenon is demonstrated during the cell cycle. In the cell cycle, Paraspeckles are present during interphase and during all of mitosis except for telophase because, when the two daughter nuclei are formed, there is no RNA Pol II transcription so the protein components instead form a perinucleolar cap. The localization patterns were also duplicated in experiments using transcription inhibiting drugs.[4]
Function
The role of the Paraspeckle is as of yet not fully understood. It has been suggested that the activity of NONO (a.k.a p54nrb), a protein component, is dependent on its localisation within the nucleus.[4] Thus, one explanation of the Paraspeckle’s function is that it provides ordered localisation of its component proteins and to thereby help direct their activity. In turn, this is believed to give the Paraspeckle a regulatory function over transcription.[10] Also, a meta-analysis by Fox et.al. (2018)[9] links the Paraspeckle’s regulation to its ability to sequester or steal component proteins and RNAs. This causes other nuclear compartments to be depleted. The current research into the Paraspeckle’s function is mainly targeted towards the roles of several of its components to indicate larger cellular use, this page mainly focuses on the roles of Paraspeckle proteins and NEAT1.
Physiological
The main insight into their physiological function is their location. Prominent Paraspeckles are only found in a subpopulation of cells in murine tissues, e.g. luteal cells or cells at the tip of the gut epithelium.[9] Hence, based on location Paraspeckles are thought to play a role in cancer regulation, reproduction and viral management.
One focus has been the Paraspeckle’s role in cancer and cell stress scenarios. Wang, Z, Li K, Huang, W (2019),[11] records that quantities of NEAT1 and thus Paraspeckles are increased in digestive system tumours and respiratory cancers. Furthermore, that expression of NEAT1 is associated with tumour size, stage of cancer, ability to spread and overall patient survival. Whilst, failure to regulate NEAT1 production has been linked to non-cancerous diseases, such as neurodegenerative diseases like Parkinson’s or Alzheimer’s.[11] However, the function of NEAT1 and Paraspeckles is not always positive, it has been proven that they enhance the malignancy and stemness of breast tumours by increasing the expression of the WNT4 gene.[4]
NEAT1 also affects pregnancy and fertility chances, especially in female mammals whose luteal cells are regulated by Paraspeckles.[10] This can cause malformation or potential no formation of the corpus luteum leading to infertility, smaller litters, and fewer viable pregnancies. In a study by Chai Y, Liu J, Zhang Z, Liu L (2016),[9] knockout mice (no NEAT1) exhibited malfunctions in epithelial cell proliferation, causing mothers to lactate poorly and reduced litter survival even further. Interestingly these knockout mice exhibit the stochastic effect; the corpus luteum will form in some, but not in all.[9] This reinforces the fact that Paraspeckles are inducible by cell stress and that environmental triggers have an impact.
From a viral aspect, NEAT1 levels have an observable impact on infections within cells with many different RNA viruses, including Japanese encephalitis, rabies,[11] HIV,[11][12] influenza,[13] and Hantaan,[14] as well as the DNA-encoded herpes simplex virus.[15] Wang, Z, Li K, Huang, W (2019)[11] suggest NEAT1_2/Paraspeckles act as a promoter to cell defence triggering and aiding the cellular defence mechanism.
Molecular
From the molecular perspective, this page examines the Paraspeckle’s function through NEAT1, NONO (p45nrb) and SFPQ (PSF).
One aspect of the Molecular function is the Paraspeckle’s ability to sequester other molecules affecting transcription. This is done by both NEAT1 and some constituent proteins. NEAT1 is primarily responsible for the Paraspeckle's architecture and providing stability to the protein components.[11] Yet, it has also been shown to regulate gene expression. This is done by recruiting transcription factors, sequestering them from gene promoters and ultimately altering transcription.[5][16][17] Furthermore, Wang, Z, Li K, Huang, W (2019)[11] state that NEAT1 can regulate expression by associating with RNA-binding proteins this regulates RNA splicing events and can manipulate the stability of proteins. Another form of molecule sequestering is through NONO and SFPQ, both proteins can bind to double-stranded RNA that has formed as a result of transcribed inverted repeat motifs.[9]
Another aspect of molecular function is NEAT1’s localisation of Paraspeckle proteins to direct their activity. In a study by Hirose, T. et al. (2014),[6] when NEAT1_2 levels increase, Paraspeckles elongate. This, in turn, not only increases Paraspeckle length but also the demand for more Paraspeckle proteins to build the tertiary structures required for proper functioning. This reduces nucleoplasmic protein availability. It was noted in their study that this has an impact on the role of free Paraspeckle proteins such as SFPQ which normally represses IL-8, an immune-responsive gene, or can activate the ADARB2 gene. Thus, gene regulation can be manipulated not just through sequestering of non-constituent proteins but also Paraspeckle constitutive proteins.
Paraspeckle Composition
| Gene Name | Importance in paraspeckle formation | Prionlike domain(a) | ALS Mutation(b) | Liquid-Liquid phase separation link(c) | Paraspeckle Zone(d) |
| PARASPECKLE PROTEINS | |||||
|---|---|---|---|---|---|
| AHDC1 | Dispensable | No | |||
| AKAP8L | Dispensable | Yes | |||
| CELF6 | n.d.(e) | No | |||
| CIRBP | Dispensable | No | |||
| CPSF5 | Dispensable | No | |||
| CPSF6 | Dispensable | No | |||
| CPSF7 | Important | No | |||
| DAZAP1 | Essential | Yes | |||
| DLX3 | n.d. | Yes | |||
| EWSR1 | Dispensable | Yes | Yes | ||
| FAM98A | Important | Yes | |||
| FIGN | Important | Yes | |||
| FUS | Essential | Yes | Yes | Yes | Core |
| HNRNPA1 | Important | Yes | Yes | Yes | |
| HNRNPA1L2 | n.d. | Yes | |||
| HNRNPF | n.d. | No | |||
| HNRNPH1 | n.d. | Yes | |||
| HNRNPH3 | Essential | Yes | |||
| HNRNPK | Essential | No | |||
| HNRNPR | Important | Yes | |||
| HNRNPUL1 | Important | Yes | |||
| MEX3A | n.d. | No | |||
| NONO | Essential | Yes | Core | ||
| PCED1A | Important | No | |||
| PSPC1 | Dispensable | Yes | Core | ||
| RBM3 | Dispensable | Yes | |||
| RBM4B | Dispensable | No | |||
| RBM7 | Dispensable | No | |||
| RBM12 | Important | Yes | |||
| RBM14 | Essential | Yes | Yes | Patch | |
| RBMX | Dispensable | No | |||
| RUNX3 | Dispensable | Yes | |||
| SFPQ | Essential | Yes | Yes | Core | |
| SMARCA4 (BRG1) | Essential | No | Patch | ||
| SRSF10 | Important | No | |||
| SS18L1 | n.d. | Yes | Yes | ||
| TAF14 | Important | Yes | Yes | ||
| TDP43 | n.d. | Yes | Yes | Shell | |
| UBAP2L | Dispensable | Yes | |||
| ZC3HG | Dispensable | Yes | |||
| PARASPECKLE RNAs | |||||
| NEAT1 | Essential | N/A(e) | 5' + 3' Shell, middle core | ||
| IR-containing RNAs | Dispensable | N/A | |||
| AG-rich RNAs | Dispensable | N/A | Shell | ||
| (a) A type of low complexity domain rich in polar and small amino acids (Gly, Ala, Ser, Pro, Asn, Gln, Tyr) implicated in forming fibrillar higher-order aggregates
(b) Amyotrophic lateral sclerosis, also known as motor neuron disease (c) Partition of components of molecular mixtures into distinct demixed phases (e.g., oil and water). In the cell, many membrane-less organelle display liquid behaviours suggesting that they are demixed liquids (d) The paraspeckle zones relate to the super-resolution imaging of paraspeckles (e) Abbreviations: n.d., not determined; N/A, not applicable | |||||
References
- Fox A (2007-03-07). "Paraspeckle Size" (Interview). Interviewed by Sundby R. E-mail Correspondence.
- Fox AH, Lam YW, Leung AK, Lyon CE, Andersen J, Mann M, Lamond AI (January 2002). "Paraspeckles: a novel nuclear domain". Current Biology. 12 (1): 13–25. doi:10.1016/S0960-9822(01)00632-7. PMID 11790299. Archived from the original on 2012-12-08.
- Fox A, Bickmore W (2004). "Nuclear Compartments: Paraspeckles". Archived from the original on 2 May 2006. Retrieved 6 March 2007.
- Fox AH, Bond CS, Lamond AI (November 2005). "P54nrb forms a heterodimer with PSP1 that localizes to paraspeckles in an RNA-dependent manner". Molecular Biology of the Cell. 16 (11): 5304–15. doi:10.1091/mbc.E05-06-0587. PMC 1266428. PMID 16148043.
- Imamura K, Imamachi N, Akizuki G, Kumakura M, Kawaguchi A, Nagata K, et al. (February 2014). "Long noncoding RNA NEAT1-dependent SFPQ relocation from promoter region to paraspeckle mediates IL8 expression upon immune stimuli". Molecular Cell. 53 (3): 393–406. doi:10.1016/j.molcel.2014.06.013. PMID 24507715.
- Hirose T, Virnicchi G, Tanigawa A, Naganuma T, Li R, Kimura H, et al. (January 2014). "NEAT1 long noncoding RNA regulates transcription via protein sequestration within subnuclear bodies". Molecular Biology of the Cell. 25 (1): 169–83. doi:10.1091/mbc.e13-09-0558. PMC 3873887. PMID 24173718.
- West JA, Mito M, Kurosaka S, Takumi T, Tanegashima C, Chujo T, et al. (September 2016). "Structural, super-resolution microscopy analysis of paraspeckle nuclear body organization". The Journal of Cell Biology. 214 (7): 817–30. doi:10.1083/jcb.201601071. PMC 5037409. PMID 27646274.
- Hu SB, Yao RW, Chen LL (September 2016). "Shedding light on paraspeckle structure by super-resolution microscopy". The Journal of Cell Biology. 214 (7): 789–91. doi:10.1083/jcb.201609008. PMC 5037413. PMID 27646270.
- Fox AH, Nakagawa S, Hirose T, Bond CS (February 2018). "Paraspeckles: Where Long Noncoding RNA Meets Phase Separation" (PDF). Trends in Biochemical Sciences. 43 (2): 124–135. doi:10.1016/j.tibs.2017.12.001. PMID 29289458.
- Schuldt A (February 2002). "Proteomics of the nucleolus". Nature Cell Biology. 4 (2): E35. doi:10.1038/ncb0202-e35. PMID 11835055. S2CID 12179853.
- Wang Z, Li K, Huang W (March 2020). "Long non-coding RNA NEAT1-centric gene regulation". Cellular and Molecular Life Sciences. doi:10.1007/s00018-020-03503-0. PMID 32219465. S2CID 214647692.
- Piskounova E, Polytarchou C, Thornton JE, LaPierre RJ, Pothoulakis C, Hagan JP, et al. (November 2011). "Lin28A and Lin28B inhibit let-7 microRNA biogenesis by distinct mechanisms". Cell. 147 (5): 1066–79. doi:10.1016/j.cell.2011.10.039. PMC 3227872. PMID 22118463.
- Llères D, Denegri M, Biggiogera M, Ajuh P, Lamond AI (June 2010). "Direct interaction between hnRNP-M and CDC5L/PLRG1 proteins affects alternative splice site choice". EMBO Reports. 11 (6): 445–51. doi:10.1038/embor.2010.64. PMC 2892320. PMID 20467437.
- Yong W, Yu D, Jun Z, Yachen D, Weiwei W, Midie X, et al. (August 2018). "Long noncoding RNA NEAT1, regulated by LIN28B, promotes cell proliferation and migration through sponging miR-506 in high-grade serous ovarian cancer". Cell Death & Disease. 9 (9): 861. doi:10.1038/s41419-018-0908-z. PMC 6113267. PMID 30154460.
- Wang Z, Li K, Wang X, Huang W (May 2019). "MiR-155-5p modulates HSV-1 replication via the epigenetic regulation of SRSF2 gene expression". Epigenetics. 14 (5): 494–503. doi:10.1080/15592294.2019.1600388. PMC 6557561. PMID 30950329.
- Wang Z, Fan P, Zhao Y, Zhang S, Lu J, Xie W, et al. (March 2017). "NEAT1 modulates herpes simplex virus-1 replication by regulating viral gene transcription". Cellular and Molecular Life Sciences. 74 (6): 1117–1131. doi:10.1007/s00018-016-2398-4. PMC 5309293. PMID 27783096.
- Wang Z, Zhao Y, Xu N, Zhang S, Wang S, Mao Y, et al. (August 2019). "NEAT1 regulates neuroglial cell mediating Aβ clearance via the epigenetic regulation of endocytosis-related genes expression". Cellular and Molecular Life Sciences. 76 (15): 3005–3018. doi:10.1007/s00018-019-03074-9. PMC 6647258. PMID 31006037.
External links
- The Nuclear Compartments:Paraspeckle page on the Nuclear Protein Database, written by Dr. Archa Fox and Dr. Wendy Bickmore, provides a factsheet and links to information on paraspeckle components.