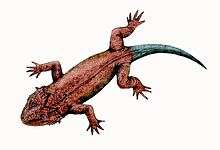
Mesosaur
Mesosaurs ("middle lizards") were a group of small aquatic reptiles that lived during the early Permian period, roughly 299 to 270 million years ago. Mesosaurs were the first known aquatic reptiles, having apparently returned to an aquatic lifestyle from more terrestrial ancestors. Exactly how terrestrial mesosaur ancestors had become remains uncertain; recent research cannot establish with confidence if the first amniotes were fully terrestrial, or only amphibious.[1] Most authors consider mesosaurs to have been aquatic,[1][2] although adult animals may have been amphibious, rather than completely aquatic, as indicated by their moderate skeletal adaptations to a semiaquatic lifestyle.[3][4] Similarly, their affinities are uncertain; they may be either the most basal sauropsids,[5][6] or among the most basal parareptiles.[7][8]
| Mesosaurs | |
|---|---|
 | |
| Mesosaurus | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | †Parareptilia |
| Order: | †Mesosauria Seeley, 1892 |
| Family: | †Mesosauridae Baur, 1889 |
| Genera | |
Phylogeny
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
The phylogenetic position of mesosaurs has an important bearing on the definition of Reptilia. In one of the first major phylogenetic studies of amniotes (vertebrates laying eggs on land) Gauthier et al. (1988) placed Mesosauridae in a group called Parareptilia.[10] Parareptilia means "at the side of reptiles" and was placed outside the clade Reptilia, which was considered a crown group. As a crown group, Reptilia included the most recent common ancestor of the two main lineages of living reptiles -diapsids and turtles- and all descendants of that common ancestor. In this sense, Reptilia was a node-based taxon because the first reptilian common ancestor would have been a "node" on the phylogenetic tree. Under this phylogeny, many extinct forms traditionally regarded as reptiles including mesosaurs were excluded from the group because they were outside the node.[9]

The study of Laurin and Reisz (1995) was the second major phylogenetic analysis of amniotes.[11] Like Gauthier et al., Laurin and Reisz used Reptilia as a crown group and placed mesosaurs outside the group. Their phylogeny differed in that the parareptiles of Gauthier et al. were now regarded as close relatives of turtles, within crown group Reptilia. Laurin and Reisz adopted the name Sauropsida as a node-based taxon including the last common ancestor of mesosaurs and Reptilia. Traditionally, amniotes are divided into two groups: a mammal lineage called Synapsida and a reptile lineage called either Reptilia or Sauropsida. In fact, the study of Gauthier (1994) defined Sauropsida as all amniotes more closely related to reptiles than to mammals, which meant that Sauropsida was a stem-based taxon encompassing the entire reptilian lineage or reptilian "stem" of Amniota (Synapsida was the mammalian stem). Under this phylogeny, the only group that prevents Sauropsida from being equivalent to Reptilia is mesosaurs.[9]
More recent phylogenetic analyses, such as that of Modesto (1999), support that of Gauthier et al. (1988) by placing mesosaurs with parareptiles.[7] However, these phylogenies follow Laurin and Reisz (1995) in placing Parareptilia within crown-group Reptilia, meaning that mesosaurs are once again members of Reptilia. Using Laurin and Reisz's node-based definition of Sauropsida as "The last common ancestor of mesosaurs, testudines and diapsids, and all its descendants",[11] Sauropsida and Reptilia are equivalent groupings; mesosaurs and testudines are more closely related to each other than either group is to diapsids,[a] meaning that the clade containing testudines and diapsids (which would be crown-group Reptilia) must also contain mesosaurs. Since Reptilia was named earlier than Sauropsida, it is used most often in modern phylogenetic analyses.[9]
A 2017 phylogenetic analysis by Laurin (who had previously published the 1995 study) and Piñeiro recovered mesosaurs as a basal member of Sauropsida/Reptilia and no longer present within Parareptilia, with Parareptilia being redefined as including former members of Procolophonomorpha (found to be paraphyletic), Millerosauria, Pareiasauria, and Pantestudines, with the latter two being found to be sister groups to one another. Parareptilia was also found to actually nest inside Diapsida as the sister group to Neodiapsida.[13]
In 2012 it was revealed that Mesosaurus has holes at the back of the skull called lower temporal fenestrae, a characteristic once thought to be present only in synapsids and diapsids.[14] The condition in the skull of Mesosaurus is most similar to that in synapsid skulls because both lack the upper temporal fenestrae of diapsids. Lower temporal fenestrae are so far known only in Mesosaurus, but may be present in all mesosaurs. The presence or absence of temporal fenestrae is an important consideration in the phylogeny of mesosaurs and other amniotes because the three major groups of amniotes -Synapsida, Diapsida, and Anapsida- have been named after the number of holes in their skull; Diapsida means "two arches" in reference to the two bars that close off the upper and lower fenestra, Synapsida means "fused arch" in reference to a single bar at the bottom of the skull closing a single fenestra, and Anapsida means "no arch" in reference to skulls that lack any bars or fenestrae. Mesosaurs were traditionally classified as anapsids because they were thought to have lacked temporal fenestrae. However, the occurrence of fenestrae in amniotes has been recognized a highly variable feature within the group for many years prior to their discovery in Mesosaurus; many anapsids such as Candelaria, Bolosaurus, and lanthanosuchoids possess lower temporal fenestrae. Furthermore, Anapsida is rarely considered a valid clade in recent phylogenetic analyses.
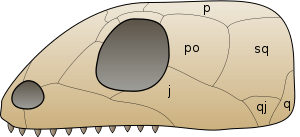
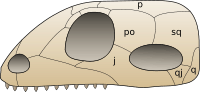
The phylogenetic position of mesosaurs influences the current understanding of how amniotes evolved temporal fenestrae. If the phylogeny produced by Laurin and Reisz (1995) is correct in that mesosaurs are basal sauropsids, the lower temporal fenestra may be a primitive feature in amniotes, present in amniote's most recent common ancestor. Synapsids would have retained their fenestrae, and so too would sauropsids except for turtles and most parareptiles. Another possibility under Laurin and Reisz's phylogeny is that lower temporal fenestrae evolved independently in mesosaurs, synapsids, diapsids, and some parareptiles, and that the lack of fenestrae is a primitive feature in amniotes. If instead mesosaurs are members of Parareptilia, the presence of temporal fenestrae is probably not a primitive feature in amniotes, and the lower temporal fenestrae in mesosaurs may be characteristic of a lineage of basal parareptiles that also includes fenestra-bearing lanthanosuchoids and Bolosaurus.[14]
Biology
They have long been thought to have been coastal forms that probably inhabited relatively shallow water,[15] but recent research suggests that at least those from Uruguay inhabited a hypersaline environment, rather than a coastal marine environment.[16] Recently described embryos show that pachyostosis of the ribs (which were thicker and denser than in terrestrial tetrapods) developed even before hatching, which suggests that mesosaurs were able to swim at birth, or shortly thereafter. They were apparently not very fast swimmers, with an optimal swimming speed estimated to have been between 0.15 and 0.86 m/s, but this must have been somewhat faster than the speed of their main prey, the pygocephalomorph crustaceans.[17] Their reproductive mode is somewhat uncertain because association between adults and possible embryos in utero suggests viviparity, as in many aquatic reptiles, but a potentially isolated egg has also been found.[18]
Recently, evidence of predation on both pygocephalomorph crustaceans and members of their own species has been established. It is thought that mesosaurs were in general adapted to hypersaline habitats.[19]
A study on the vertebral column torso and tail proportions of Mesosaurus suggests that, while juveniles may have been fully aquatic, adults might have spent some time on land; this is further vindicated by the rarity of adult animals in aquatic settings and some faeces showing signs of drying fracture. However, how terrestrial they were is difficult to say, as this same study states that terrestrial foraging would have been difficult due to their speciations to an aquatic life.[20]
Notes
- ^ Studies using molecular phylogenetics, which examine the genes and proteins of living organisms, suggest that testudines (turtles) are diapsids. These studies show that mesosaurs do not form a clade with turtles that excludes diapsids, but fossil evidence still suggests that mesosaurs form a group with parareptiles. In most recent studies, Reptilia is not used as a crown group and still contains mesosaurs and Parareptilia.
References
- Canoville, A. and M. Laurin. 2010. Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on paleobiological inferences. Biological Journal of the Linnean Society 100:384–406.
- Modesto, S. 2006. The cranial skeleton of the Early Permian aquatic reptile Mesosaurus tenuidens: implications for relationships and palaeobiology. Zoological Journal of the Linnean Society 146:345-368.
- Piñeiro, G. 2008. Los mesosaurios y otros fosiles de fines del Paleozoico; pp. 179–205 in D. Perera (ed.), Fósiles de Uruguay. DIRAC, Montevideo.
- Pablo Nuñez Demarco et al. Was Mesosaurus a Fully Aquatic Reptile? Front. Ecol. Evol, published online July 27, 2018; doi: 10.3389/fevo.2018.00109
- Laurin, M. and R. R. Reisz. 1995. A reevaluation of early amniote phylogeny. Zoological Journal of the Linnean Society 113:165-223.
- Laurin, Michel; Piñeiro, Graciela H. (2017). "A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes". Frontiers in Earth Science. 5. doi:10.3389/feart.2017.00088.
- Modesto, S.P. (1999). "Observations of the structure of the Early Permian reptile Stereosternum tumidum Cope". Palaeontologia Africana. 35: 7–19.
- MacDougall, M.J.; Modesto, S.P.; Brocklehurst,N.; Verriere, A.; Reisz, R.R.; Fröbisch, J. (2018). "Response: A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes". Front. Earth Sci. 6:99 doi: 10.3389/feart.2018.00099
- Modesto, S.P.; Anderson, J.S. (2004). "The phylogenetic definition of Reptilia" (PDF). Systematic Biology. 53 (5): 815–821. doi:10.1080/10635150490503026. PMID 15545258. Archived from the original (PDF) on 2016-07-05. Retrieved 2012-05-19.
- Gauthier, J.A.; Kluge, A.G.; Rowe, T. (1988). "The early evolution of the Amniota". In Benton, M.J. (ed.). The Phylogeny and Classification of the Tetrapods. 1. Oxford: Clarendon Press. pp. 103–155. ISBN 978-0198577058.
- Laurin, M.; Reisz, R.R. (1995). "A reevaluation of early amniote phylogeny" (PDF). Zoological Journal of the Linnean Society. 113 (2): 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x.
- Laurin, Michel; Piñeiro, Graciela H. (2017). "A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes". Frontiers in Earth Science. 5. doi:10.3389/feart.2017.00088. ISSN 2296-6463.
- Laurin, Michel; Piñeiro, Graciela H. (2017). "A Reassessment of the Taxonomic Position of Mesosaurs, and a Surprising Phylogeny of Early Amniotes". Frontiers in Earth Science. 5. doi:10.3389/feart.2017.00088. ISSN 2296-6463.
- Piñeiro, G.; Ferigolo, J.; Ramos, A.; Laurin, M. (2012). "Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed". Comptes Rendus Palevol. 11 (5): 379–391. doi:10.1016/j.crpv.2012.02.001.
- Oelofsen, B. and D. C. Araújo. 1983. Palaeoecological implications of the distribution of mesosaurid reptiles in the Permian Irati sea (Paraná basin), South America. Revista Brasileira de Geociências 13:1–6.
- Piñeiro, G.; Ramos, A.; Goso, C.; Scarabino, F.; Laurin, M. (2012). "Unusual environmental conditions preserve a Permian mesosaur-bearing Konservat-Lagerstätte from Uruguay" (PDF). Acta Palaeontologica Polonica. 57 (2): 299–318. doi:10.4202/app.2010.0113.
- Villamil, J. N.; Demarco, P. N.; Meneghel, M.; Blanco, R. E.; Jones, W.; Rinderknecht, A. S.; Laurin, M.; Pineiro, G. (2015). "Optimal swimming speed estimates in the Early Permian mesosaurid Mesosaurus tenuidens (Gervais 1867) from Uruguay". Historical Biology. 28 (7): 963–971. doi:10.1080/08912963.2015.1075018.
- Piñeiro, G.; Ferigolo, J.; Meneghel, M.; Laurin, M. (2012). "The oldest known amniotic embryos suggest viviparity in mesosaurs". Historical Biology. 24 (6): 620–630. doi:10.1080/08912963.2012.662230.
- Rivaldo R. Da Silva; Jorge Ferigolo; Piotr Bajdek; Graciela H. Piñeiro (2017). "The feeding habits of Mesosauridae". Frontiers in Earth Science. 5: Article 23. doi:10.3389/feart.2017.00023.
- Pablo Nuñez Demarco et al. Was Mesosaurus a Fully Aquatic Reptile? Front. Ecol. Evol, published online July 27, 2018; doi: 10.3389/fevo.2018.00109