Leucyl aminopeptidase
Leucyl aminopeptidases (EC 3.4.11.1, leucine aminopeptidase, LAPs, leucyl peptidase, peptidase S, cytosol aminopeptidase, cathepsin III, L-leucine aminopeptidase, leucinaminopeptidase, leucinamide aminopeptidase, FTBL proteins, proteinates FTBL, aminopeptidase II, aminopeptidase III, aminopeptidase I) are enzymes that preferentially catalyze the hydrolysis of leucine residues at the N-terminus of peptides and proteins. Other N-terminal residues can also be cleaved, however. LAPs have been found across superkingdoms. Identified LAPs include human LAP, bovine lens LAP, porcine LAP, Escherichia coli (E. coli) LAP (also known as PepA or XerB), and the solanaceous-specific acidic LAP (LAP-A) in tomato (Solanum lycopersicum).
| Leucine aminopeptidase | |
|---|---|
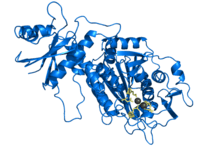 Crystal structure of bovine leucyl aminopeptidase with co-ordinated zinc ions. Rendered from PDB 1BLL. | |
| Identifiers | |
| Symbol | LAP |
| Alt. symbols | PEPS |
| NCBI gene | 51056 |
| HGNC | 18449 |
| OMIM | 170250 |
| RefSeq | NM_015907 |
| UniProt | P28838 |
| Other data | |
| EC number | 3.4.11.1 |
| Locus | Chr. 4 p15.33 |
Enzyme description, structure, and active site
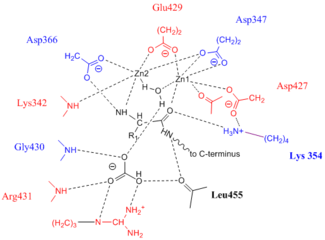
The active sites in PepA and in bovine lens LAP have been found to be similar.[1] Shown in the picture below is the proposed model for the active site of LAP-A in tomato based on the work of Strater et al.[2][3] It is also known that the biochemistry of the LAPs from these three kingdoms is very similar. PepA, bovine lens LAP, and LAP-A preferentially cleave N-terminal leucine, arginine, and methionine residues. These enzymes are all metallopeptidases requiring divalent metal cations for their enzymatic activity[4] Enzymes are active in the presence of Mn+2, Mg+2 and Zn+2. These enzymes are also known to have high pH (pH 8) and temperature optima. At pH 8, the highest enzymatic activity is seen at 60 °C. PepA, bovine lens LAP and LAP-A are also known to form hexamers in vivo. The Gu et al. from 1999 demonstrated that six 55kDA enzymatically inactive LAP-A protomers come together to form the 353kDa bioactive LAP-A hexamer. Structures of the bovine lens LAP protomer and the biologically active hexamer have been constructed[5] can be found through Protein Data Bank (2J9A).
Mechanism(s)
Historically, the mechanisms of carboxypeptidases and endoprotease have been much more well-studied and understood by researchers (Ref #6 Lipscomb 1990). Work within the past two decades has provided vital knowledge regarding the mechanisms of aminopeptidases. The mechanism of
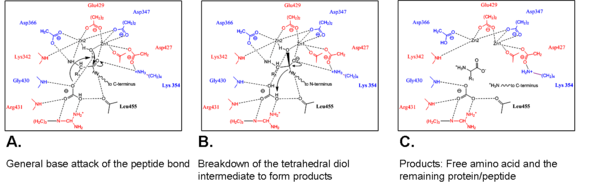
bovine lens LAP and PepA have been elucidated (Ref 1 and 2), however, the exact mechanism of tomato LAP-A is unknown at this time. A search of current literature does not indicate that new research is underway to determine the exact mechanism of LAP-A. Based on the biochemical similarities of the LAPs between kingdoms, the mechanism of LAP-A may be similar to bovine lens LAP and PepA.
Biological function
Once thought of as a housekeeping gene necessary only for protein turnover, studies have demonstrated that LAP-A has a regulatory role in the immune response in tomato.
Background on plant immune response
In order to survive, plants must be able to respond to many biotic and abiotic stresses, including pathogen attack, piercing/sucking insects, herbivory, and mechanical wounding. These stresses activate specialized signal transduction pathways, which are specific to the stressor and the amount of tissue damage inflicted. Similar to mechanical wounding, chewing insects, such as the tobacco hornworm (Manduca sexta, one of the major pests of tomato), cause extensive tissue damage activating the jasmonic acid (JA)-mediated response (Walling 2000). This JA-mediated response revolves around the octadecanoid pathway, which is responsible for the synthesis of JA and several other potent signaling molecules, and ends in the regulation of two sets of genes whose expression changes over time. The early genes amplify the wounding signal and can be detected 30 minutes to 2 hours after damage (Ryan 2000). Late gene expression can be seen 4–24 hours after wounding. Products of late-response genes act as deterrents to chewing-insect feeding, often by decreasing the nutritional value of the food ingested or interfering with insect gut function (Walling 2000). For example, serine proteinase inhibitors (Pins) interfere with digestive proteases in the insect gut and polyphenol oxidases (PPO) act to decrease the nutritive value of plant leaves after ingestion by herbivores (Johnson et al. 1989; Ryan 2000; Orozco-Cardenas 2001). Please see the Picture 3 for a summary of the wound response in tomato.
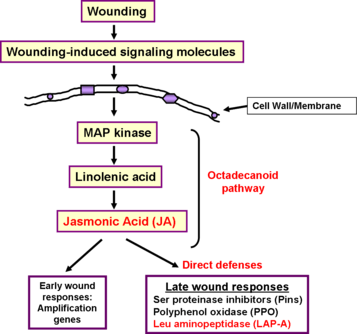
The plant response in this octadecanoid pathway is similar to mammalian prostaglandin and leukotriene pathways (Ref Walling 2000). This particular pathway is inhibited by salicylic acid.
Octadecanoid pathway
(LAP-A), a product of the octadecanoid pathway in some solanaceous plants, has been shown by Fowler et al. to have a regulatory role in the late wound response of tomato. Experiments were conducted using three genotypes of tomato plants: wildtype (WT), (LapA-SI) plants that were silenced for LAP-A, and LapA-OX that constitutively expressed LAP-A. Late-gene expression was inhibited in wounded LapA-SI plants, and the LapA-SI plants were also more susceptible to tobacco hornworm feeding, relative to wildtype (WT) plants. In comparison, the wounded LapA-OX leaves exhibited heightened levels of late gene RNA accumulation, an increased resistance to herbivory, and extended expression of late wound-response genes. These data suggest that LAP-A functions in regulating both the intensity and the persistence of the late wound response. However, unwounded LapA-OX did not accumulate late gene RNA transcripts, suggesting that presence of LAP-A alone is not sufficient to induce late gene expression. LAP-A is the first plant aminopeptidase shown to have a regulatory role in signal transduction pathway.
Osmoregulation
LAP proteins are expressed in a variety of marine organisms as a method of coping with the osmotic threat high salinity poses to the cell. During bouts of high salinity, LAP begins the catalysis of proteins in order to release amino acids into the cell in an attempt to balance the high ion concentrations in the external environment.[6]
References
- Sträter N, Sun L, Kantrowitz ER, Lipscomb WN (September 1999). "A bicarbonate ion as a general base in the mechanism of peptide hydrolysis by dizinc leucine aminopeptidase". Proceedings of the National Academy of Sciences of the United States of America. 96 (20): 11151–5. Bibcode:1999PNAS...9611151S. doi:10.1073/pnas.96.20.11151. PMC 18002. PMID 10500145.
- Gu YQ, Walling LL (March 2002). "Identification of residues critical for activity of the wound-induced leucine aminopeptidase (LAP-A) of tomato". European Journal of Biochemistry / FEBS. 269 (6): 1630–40. doi:10.1046/j.1432-1327.2002.02795.x. PMID 11895433.
- Gu YQ, Walling LL (March 2002). "Identification of residues critical for activity of the wound-induced leucine aminopeptidase (LAP-A) of tomato". European Journal of Biochemistry / FEBS. 269 (6): 1630–40. doi:10.1046/j.1432-1327.2002.02795.x. PMID 11895433.
- Gu YQ, Holzer FM, Walling LL (August 1999). "Overexpression, purification and biochemical characterization of the wound-induced leucine aminopeptidase of tomato". European Journal of Biochemistry / FEBS. 263 (3): 726–35. doi:10.1046/j.1432-1327.1999.00548.x. PMID 10469136.
- Kraft M, Schleberger C, Weckesser J, Schulz GE (December 2006). "Binding structure of the leucine aminopeptidase inhibitor microginin FR1". FEBS Letters. 580 (30): 6943–7. doi:10.1016/j.febslet.2006.11.060. PMID 17157838.
- Hilbish TJ (1985). "The Physiological Basis of Natural Selection at the Lap Locus". Evolution. 39 (6): 1302–1317. doi:10.2307/2408787. JSTOR 2408787.
- Sources
- Sträter N, Sun L, Kantrowitz ER, Lipscomb WN (September 1999). "A bicarbonate ion as a general base in the mechanism of peptide hydrolysis by dizinc leucine aminopeptidase". Proceedings of the National Academy of Sciences of the United States of America. 96 (20): 11151–5. Bibcode:1999PNAS...9611151S. doi:10.1073/pnas.96.20.11151. PMC 18002. PMID 10500145.
- Sträter N, Lipscomb WN (November 1995). "Two-metal ion mechanism of bovine lens leucine aminopeptidase: active site solvent structure and binding mode of L-leucinal, a gem-diolate transition state analogue, by X-ray crystallography". Biochemistry. 34 (45): 14792–800. doi:10.1021/bi00045a021. PMID 7578088.
- Gu YQ, Walling LL (March 2002). "Identification of residues critical for activity of the wound-induced leucine aminopeptidase (LAP-A) of tomato". European Journal of Biochemistry / FEBS. 269 (6): 1630–40. doi:10.1046/j.1432-1327.2002.02795.x. PMID 11895433.
- Gu YQ, Holzer FM, Walling LL (August 1999). "Overexpression, purification and biochemical characterization of the wound-induced leucine aminopeptidase of tomato". European Journal of Biochemistry / FEBS. 263 (3): 726–35. doi:10.1046/j.1432-1327.1999.00548.x. PMID 10469136.
- Kraft M, Schleberger C, Weckesser J, Schulz GE (December 2006). "Binding structure of the leucine aminopeptidase inhibitor microginin FR1". FEBS Letters. 580 (30): 6943–7. doi:10.1016/j.febslet.2006.11.060. PMID 17157838.
- Walling LL (June 2000). "The Myriad Plant Responses to Herbivores". Journal of Plant Growth Regulation. 19 (2): 195–216. doi:10.1007/s003440000026. PMID 11038228.
- Burley SK, David PR, Lipscomb WN (August 1991). "Leucine aminopeptidase: bestatin inhibition and a model for enzyme-catalyzed peptide hydrolysis". Proceedings of the National Academy of Sciences of the United States of America. 88 (16): 6916–20. Bibcode:1991PNAS...88.6916B. doi:10.1073/pnas.88.16.6916. PMC 52204. PMID 1871107.
- Orozco-Cárdenas ML, Narváez-Vásquez J, Ryan CA (January 2001). "Hydrogen peroxide acts as a second messenger for the induction of defense genes in tomato plants in response to wounding, systemin, and methyl jasmonate". The Plant Cell. 13 (1): 179–91. doi:10.2307/3871162. JSTOR 3871162. PMC 102208. PMID 11158538.
- Ryan CA (March 2000). "The systemin signaling pathway: differential activation of plant defensive genes". Biochimica et Biophysica Acta. 1477 (1–2): 112–21. doi:10.1016/S0167-4838(99)00269-1. PMID 10708853.
- Johnson R, Narvaez J, An G, Ryan C (December 1989). "Expression of proteinase inhibitors I and II in transgenic tobacco plants: effects on natural defense against Manduca sexta larvae". Proceedings of the National Academy of Sciences of the United States of America. 86 (24): 9871–5. Bibcode:1989PNAS...86.9871J. doi:10.1073/pnas.86.24.9871. PMC 298604. PMID 2602379.
- Fowler JH, Narváez-Vásquez J, Aromdee DN, Pautot V, Holzer FM, Walling LL (April 2009). "Leucine aminopeptidase regulates defense and wound signaling in tomato downstream of jasmonic acid". The Plant Cell. 21 (4): 1239–51. doi:10.1105/tpc.108.065029. PMC 2685619. PMID 19376935.
External links
- The MEROPS online database for peptidases and their inhibitors: Animal:M17.001, Bacteria:M17.003, Plant::M17.002.
- Leucyl+aminopeptidase at the US National Library of Medicine Medical Subject Headings (MeSH)