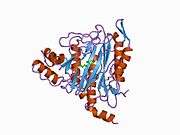
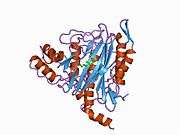
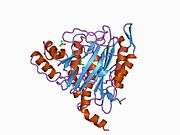
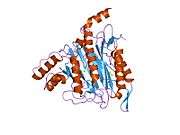
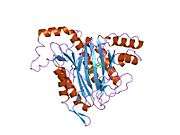
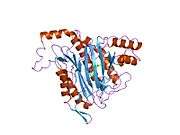
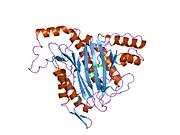
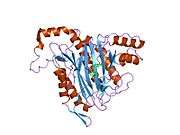
METAP2
Methionine aminopeptidase 2 is an enzyme that in humans is encoded by the METAP2 gene.[5][6]
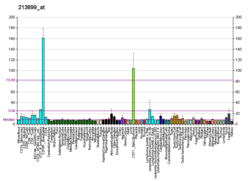
Methionine aminopeptidase 2, a member of the dimetallohydrolase family, is a cytosolic metalloenzyme that catalyzes the hydrolytic removal of N-terminal methionine residues from nascent proteins.[7][8][9]
- peptide-methionine peptide + methionine

MetAP2 is found in all organisms and is especially important because of its critical role in tissue repair and protein degradation.[7] Furthermore, MetAP2 is of particular interest because the enzyme plays a key role in angiogenesis, the growth of new blood vessels, which is necessary for the progression of diseases including solid tumor cancers and rheumatoid arthritis.[10] MetAP2 is also the target of two groups of anti-angiogenic natural products, ovalicin and fumagillin, and their analogs.[11][12][13][14]
Structure
In living organisms, the start codon that initiates protein synthesis codes for either methionine (eukaryotes) or formylmethionine (prokaryotes). In E. coli (prokaryote), an enzyme called formylmethionine deformylase can cleave the formyl group, leaving just the N-terminal methionine residue. For proteins with small, uncharged penultimate N-terminal residues, a methionine aminopeptidase can cleave the methionine residue.[7] The number of genes encoding for a methionine aminopeptidase varies between organisms. In E. coli, there is only one known MetAP, a 29,333 Da monomeric enzyme coded for by a gene consisting of 264 codons.[7] The knockout of this gene in E. coli leads to cell inviability.[15] In humans, there are two genes encoding MetAP, MetAP1 and MetAP2. MetAP1 codes for a 42 kDa enzyme, while MetAP2 codes for a 67 kDa enzyme. Yeast MetAP1 is 40 percent homologous to E. coli MetAP; within S. cerevisiae, MetAP2 is 22 percent homologous with the sequence of MetAP1; MetAP2 is highly conserved between S. cerevisiae and humans.[16] In contrast to prokaryotes, eukaryotic S. cerevisiae strains lacking the gene for either MetAP1 or MetAP2 are viable, but exhibit a slower growth rate than a control strain expressing both genes.
Active site
The active site of MetAP2 has a structural motif characteristic of many metalloenzymes—including the dioxygen carrier protein, hemerythrin; the dinuclear non-heme iron protein, ribonucleotide reductase; leucine aminopeptidase; urease; arginase; several phosphatases and phosphoesterases—that includes two bridging carboxylate ligands and a bridging water or hydroxide ligand.[7][8][17][18][19][20][21] Specifically in human MetAP2 (PDB: 1BOA), one of the catalytic metal ions is bound to His331, Glu364, Glu459, Asp263, and a bridging water or hydroxide, while the other metal ion is bound to Asp251 (bidentate), App262 (bidentate), Glu459, and the same bridging water or hydroxide. Here, the two bridging carboxylates are Asp262 and Glu459.
Dimetal center
The identity of the active site metal ions under physiological conditions has not been successfully established, and remains a controversial issue. MetAP2 shows activity in the presence of Zn(II), Co(II), Mn(II), and Fe(II) ions, and various authors have argued any given metal ion is the physiological one: some in the presence of iron,[22] others in cobalt,[23][24] others in manganese,[25] and yet others in the presence of zinc.[26] Nonetheless, the majority of crystallographers have crystallized MetAP2 either in the presence of Zn(II) or Co(II) (see PDB database).
Mechanism
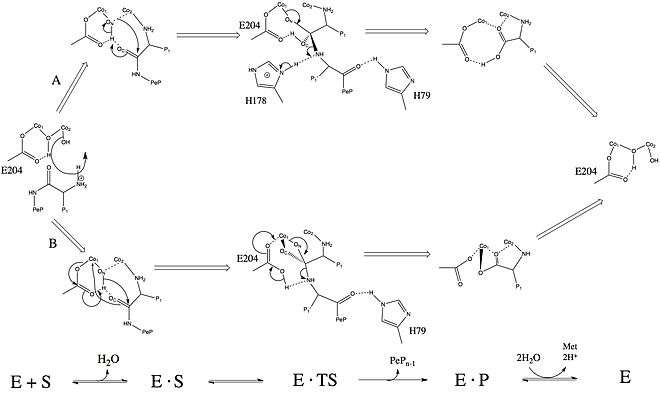
The bridging water or hydroxide ligand acts as a nucleophile during the hydrolysis reaction, but the exact mechanism of catalysis is not yet known.[10][19][28] The catalytic mechanisms of hydrolase enzymes depend greatly on the identity of the bridging ligand,[29] which can be challenging to determine due to the difficulty of studying hydrogen atoms via x-ray crystallography.
The histidine residues shown in the mechanism to the right, H178 and H79, are conserved in all MetAPs (MetAP1s and MetAP2s) sequenced to date, suggesting their presence is important to catalytic activity.[30] Based upon X-ray crystallographic data, histidine 79 (H79) has been proposed to help position the methionine residue in the active site and transfer a proton to the newly exposed N-terminal amine.[12] Lowther and Colleagues have proposed two possible mechanisms for MetAP2 in E. coli, shown at the right.[14]
Function
While previous studies have indicated MetAP2 catalyzes the removal of N-terminal methionine residues in vitro, the function of this enzyme in vivo may be more complex. For example, a significant correlation exists between the inhibition of the enzymatic activity of MetAP2 and inhibition of cell growth, thus implicating the enzyme in endothelial cell proliferation.[13] For this reason, scientists have singled out MetAP2 as a potential target for the inhibition of angiogenesis. Moreover, studies have demonstrated that MetAP2 copurifies and interacts with the α subunit of eukaryotic initiation factor 2 (eIF2), a protein that is necessary for protein synthesis in vivo.[31] Specifically, MetAP2 protects eIF-2α from inhibitory phosphorylation from the enzyme eIF-2α kinase, inhibits RNA-dependent protein kinase (PKR)-catalyzed eIF-2 R-subunit phosphorylation, and also reverses PKR-mediated inhibition of protein synthesis in intact cells.
Clinical significance
Numerous studies implicate MetAP2 in angiogenesis.[13][20][32][33][34] Specifically, the covalent binding of either the ovalicin or fumagillin epoxide moiety to the active site histidine residue of MetAP2 has been shown to inactivate the enzyme, thereby inhibiting angiogenesis. The way in which MetAP2 regulates angiogenesis has yet to be established, however, such that further study is required to validate that antiangiogenic activity results directly from MetAP2 inhibition. Nevertheless, with both the growth and metastasis of solid tumors depending heavily on angiogenesis, fumagillin and its analogs—including TNP-470, caplostatin, and beloranib—as well as ovalicin represent potential anticancer agents.[33][34] Moreover, the ability of MetAP2 to decrease cell viability in prokaryotic and small eukaryotic organisms has made it a target for antibacterial agents.[13] Thus far, both fumagillin and TNP-470 have been shown to possess antimalarial activity both in vitro and in vivo, and fumarranol, another fumagillin analog, represents a promising lead.[34]
The METAP2 inhibitor beloranib (ZGN-433) has shown efficacy in reducing weight in severely obese subjects.[35] MetAP2 inhibitors work by re-establishing balance to the ways the body metabolizes fat, leading to substantial loss of body weight. Development of beloranib was halted in 2016 after deaths during clinical trials.[36]
Interactions
METAP2 has been shown to interact with Protein kinase R.[37]
References
- GRCh38: Ensembl release 89: ENSG00000111142 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000036112 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Arfin SM, Kendall RL, Hall L, Weaver LH, Stewart AE, Matthews BW, Bradshaw RA (September 1995). "Eukaryotic methionyl aminopeptidases: two classes of cobalt-dependent enzymes". Proc Natl Acad Sci U S A. 92 (17): 7714–8. Bibcode:1995PNAS...92.7714A. doi:10.1073/pnas.92.17.7714. PMC 41216. PMID 7644482.
- Li X, Chang YH (November 1996). "Evidence that the human homologue of a rat initiation factor-2 associated protein (p67) is a methionine aminopeptidase". Biochem Biophys Res Commun. 227 (1): 152–9. doi:10.1006/bbrc.1996.1482. PMID 8858118.
- Bennett B, Holz RC (1997). "EPR Studies on the Mono- and Dicobalt(II)-Substituted Forms of the Aminopeptidase from Aeromonas proteolytica. Insight into the Catalytic Mechanism of Dinuclear Hydrolases". J. Am. Chem. Soc. 119 (8): 1923–1933. doi:10.1021/ja963021v.
- Johansson FB, Bond AD, Nielsen UG, Moubaraki B, Murray KS, Berry KJ, Larrabee JA, McKenzie CJ (June 2008). "Dicobalt II-II, II-III, and III-III complexes as spectroscopic models for dicobalt enzyme active sites". Inorg Chem. 47 (12): 5079–92. doi:10.1021/ic7020534. PMID 18494467.
- Larrabee JA, Leung CH, Moore RL, Thamrong-nawasawat T, Wessler BS (October 2004). "Magnetic circular dichroism and cobalt(II) binding equilibrium studies of Escherichia coli methionyl aminopeptidase". J. Am. Chem. Soc. 126 (39): 12316–24. doi:10.1021/ja0485006. PMID 15453765.
- Folkman J (January 1995). "Angiogenesis in cancer, vascular, rheumatoid and other disease". Nat. Med. 1 (1): 27–31. doi:10.1038/nm0195-27. PMID 7584949.
- Taunton J (July 1997). "How to starve a tumor". Chem. Biol. 4 (7): 493–6. doi:10.1016/S1074-5521(97)90320-3. PMID 9263636.
- Sin N, Meng L, Wang MQ, Wen JJ, Bornmann WG, Crews CM (June 1997). "The anti-angiogenic agent fumagillin covalently binds and inhibits the methionine aminopeptidase, MetAP-2". Proc. Natl. Acad. Sci. U.S.A. 94 (12): 6099–103. Bibcode:1997PNAS...94.6099S. doi:10.1073/pnas.94.12.6099. PMC 21008. PMID 9177176.
- Griffith EC, Su Z, Turk BE, Chen S, Chang YH, Wu Z, Biemann K, Liu JO (June 1997). "Methionine aminopeptidase (type 2) is the common target for angiogenesis inhibitors AGM-1470 and ovalicin". Chem. Biol. 4 (6): 461–71. doi:10.1016/S1074-5521(97)90198-8. PMID 9224570.
- Lowther WT, McMillen DA, Orville AM, Matthews BW (October 1998). "The anti-angiogenic agent fumagillin covalently modifies a conserved active-site histidine in the Escherichia coli methionine aminopeptidase". Proc. Natl. Acad. Sci. U.S.A. 95 (21): 12153–7. Bibcode:1998PNAS...9512153L. doi:10.1073/pnas.95.21.12153. PMC 22800. PMID 9770455.
- Chang SY, McGary EC, Chang S (July 1989). "Methionine aminopeptidase gene of Escherichia coli is essential for cell growth". J. Bacteriol. 171 (7): 4071–2. doi:10.1128/jb.171.7.4071-4072.1989. PMC 210164. PMID 2544569.
- Li X, Chang YH (December 1995). "Amino-terminal protein processing in Saccharomyces cerevisiae is an essential function that requires two distinct methionine aminopeptidases". Proc. Natl. Acad. Sci. U.S.A. 92 (26): 12357–61. Bibcode:1995PNAS...9212357L. doi:10.1073/pnas.92.26.12357. PMC 40356. PMID 8618900.
- Mizoguchi TJ, Kuzelka J, Spingler B, DuBois JL, Davydov RM, Hedman B, Hodgson KO, Lippard SJ (August 2001). "Synthesis and spectroscopic studies of non-heme diiron(III) species with a terminal hydroperoxide ligand: models for hemerythrin". Inorg Chem. 40 (18): 4662–73. doi:10.1021/ic010076b. PMID 11511213.
- Hagen KS, Lachicotte R, Kitaygorodskiy A (1993). "Supramolecular Control of Stepwise and Selective Carboxylate Ligand Substitution in Aqua-Carboxylato-Bridged Dimetal(II) Complexes". J. Am. Chem. Soc. 115 (26): 12617–12618. doi:10.1021/ja00079a064.
- Brown DA, Errington W, Glass WK, Haase W, Kemp TJ, Nimir H, Ostrovsky SM, Werner R (November 2001). "Magnetic, spectroscopic, and structural studies of dicobalt hydroxamates and model hydrolases". Inorg Chem. 40 (23): 5962–71. doi:10.1021/ic0103345. PMID 11681912.
- Larrabee JA, Chyun SA, Volwiler AS (November 2008). "Magnetic circular dichroism study of a dicobalt(II) methionine aminopeptidase/fumagillin complex and dicobalt II-II and II-III model complexes". Inorg Chem. 47 (22): 10499–508. doi:10.1021/ic8011553. PMID 18921993.
- Wilcox DE (November 1996). "Binuclear Metallohydrolases". Chem. Rev. 96 (7): 2435–2458. doi:10.1021/cr950043b. PMID 11848832.
- D'souza VM, Holz RC (August 1999). "The methionyl aminopeptidase from Escherichia coli can function as an iron(II) enzyme". Biochemistry. 38 (34): 11079–85. doi:10.1021/bi990872h. PMID 10460163.
- Chang YH, Teichert U, Smith JA (April 1992). "Molecular cloning, sequencing, deletion, and overexpression of a methionine aminopeptidase gene from Saccharomyces cerevisiae". J. Biol. Chem. 267 (12): 8007–11. PMID 1569059.
- Ghosh M, Grunden AM, Dunn DM, Weiss R, Adams MW (September 1998). "Characterization of native and recombinant forms of an unusual cobalt-dependent proline dipeptidase (prolidase) from the hyperthermophilic archaeon Pyrococcus furiosus". J. Bacteriol. 180 (18): 4781–9. doi:10.1128/JB.180.18.4781-4789.1998. PMC 107500. PMID 9733678.
- Wang J, Sheppard GS, Lou P, Kawai M, Park C, Egan DA, Schneider A, Bouska J, Lesniewski R, Henkin J (May 2003). "Physiologically relevant metal cofactor for methionine aminopeptidase-2 is manganese". Biochemistry. 42 (17): 5035–42. doi:10.1021/bi020670c. PMID 12718546.
- Leopoldini M, Russo N, Toscano M (June 2007). "Which one among Zn(II), Co(II), Mn(II), and Fe(II) is the most efficient ion for the methionine aminopeptidase catalyzed reaction?". J. Am. Chem. Soc. 129 (25): 7776–84. doi:10.1021/ja068168t. PMID 17523636.
- Lowther WT, Zhang Y, Sampson PB, Honek JF, Matthews BW (November 1999). "Insights into the mechanism of Escherichia coli methionine aminopeptidase from the structural analysis of reaction products and phosphorus-based transition-state analogues". Biochemistry. 38 (45): 14810–9. doi:10.1021/bi991711g. PMID 10555963.
- Schultz BE, Ye B, Li X, Chan SI (1997). "Electronic Paramagnetic Resonance and Magnetic Properties of Model Complexes for Binuclear Active Sites in Hydrolase Enzymes". Inorg. Chem. 36 (12): 2617–2622. doi:10.1021/ic960988r.
- Korendovych IV, Kryatov SV, Reiff WM, Rybak-Akimova EV (November 2005). "Diiron(II) mu-aqua-mu-hydroxo model for non-heme iron sites in proteins". Inorg Chem. 44 (24): 8656–8. doi:10.1021/ic051739i. PMID 16296818.
- Li JY, Cui YM, Chen LL, Gu M, Li J, Nan FJ, Ye QZ (May 2004). "Mutations at the S1 sites of methionine aminopeptidases from Escherichia coli and Homo sapiens reveal the residues critical for substrate specificity". J. Biol. Chem. 279 (20): 21128–34. doi:10.1074/jbc.M401679200. PMID 14976199.
- Wu S, Rehemtulla A, Gupta NK, Kaufman RJ (June 1996). "A eukaryotic translation initiation factor 2-associated 67 kDa glycoprotein partially reverses protein synthesis inhibition by activated double-stranded RNA-dependent protein kinase in intact cells". Biochemistry. 35 (25): 8275–80. doi:10.1021/bi953028+. PMID 8679583.
- Benny O, Fainaru O, Adini A, Cassiola F, Bazinet L, Adini I, Pravda E, Nahmias Y, Koirala S, Corfas G, D'Amato RJ, Folkman J (July 2008). "An orally delivered small-molecule formulation with antiangiogenic and anticancer activity". Nat. Biotechnol. 26 (7): 799–807. doi:10.1038/nbt1415. PMC 2803109. PMID 18587385.
- Sato Y (2004). "Aminopeptidases in Health and Disease: Role of Aminopeptidase in Angiogenesis". Biol. Pharm. Bull. 27 (6): 772–776. doi:10.1248/bpb.27.772. PMID 15187415.
- Chen X, Xie S, Bhat S, Kumar N, Shapiro TA, Liu JO (February 2009). "Fumagillin and fumarranol interact with P. falciparum methionine aminopeptidase 2 and inhibit malaria parasite growth in vitro and in vivo". Chem. Biol. 16 (2): 193–202. doi:10.1016/j.chembiol.2009.01.006. PMID 19246010.
- "Zafgen Announces Positive Topline Phase 1b Data for ZGN-433 in Obesity". MedNews. Drugs.com. 2011-01-01. Retrieved 2011-04-13.
- "Zafgen Halts Development of Beloranib, to Cut Jobs by ~34%". nasdaq.com. July 20, 2016.
- Gil J, Esteban M, Roth D (December 2000). "In vivo regulation of the dsRNA-dependent protein kinase PKR by the cellular glycoprotein p67". Biochemistry. 39 (51): 16016–25. doi:10.1021/bi001754t. PMID 11123929.
Further reading
- Prigmore E, Ahmed S, Best A, Kozma R, Manser E, Segal AW, Lim L (May 1995). "A 68-kDa kinase and NADPH oxidase component p67phox are targets for Cdc42Hs and Rac1 in neutrophils". J. Biol. Chem. 270 (18): 10717–22. doi:10.1074/jbc.270.18.10717. PMID 7738010.
- Li X, Chang YH (February 1995). "Molecular cloning of a human complementary DNA encoding an initiation factor 2-associated protein (p67)". Biochim. Biophys. Acta. 1260 (3): 333–6. doi:10.1016/0167-4781(94)00227-t. PMID 7873610.
- Ray MK, Chakraborty A, Datta B, Chattopadhyay A, Saha D, Bose A, Kinzy TG, Wu S, Hileman RE, Merrick WC (May 1993). "Characteristics of the eukaryotic initiation factor 2 associated 67-kDa polypeptide". Biochemistry. 32 (19): 5151–9. doi:10.1021/bi00070a026. PMID 8098621.
- Liu S, Widom J, Kemp CW, Crews CM, Clardy J (November 1998). "Structure of human methionine aminopeptidase-2 complexed with fumagillin". Science. 282 (5392): 1324–7. Bibcode:1998Sci...282.1324L. doi:10.1126/science.282.5392.1324. PMID 9812898.
- Griffith EC, Su Z, Niwayama S, Ramsay CA, Chang YH, Liu JO (December 1998). "Molecular recognition of angiogenesis inhibitors fumagillin and ovalicin by methionine aminopeptidase 2". Proc. Natl. Acad. Sci. U.S.A. 95 (26): 15183–8. Bibcode:1998PNAS...9515183G. doi:10.1073/pnas.95.26.15183. PMC 28017. PMID 9860943.
- Datta B, Datta R, Mukherjee S, Zhang Z (1999). "Increased phosphorylation of eukaryotic initiation factor 2alpha at the G2/M boundary in human osteosarcoma cells correlates with deglycosylation of p67 and a decreased rate of protein synthesis". Exp. Cell Res. 250 (1): 223–30. doi:10.1006/excr.1999.4508. PMID 10388536.
- Gil J, Esteban M, Roth D (2001). "In vivo regulation of the dsRNA-dependent protein kinase PKR by the cellular glycoprotein p67". Biochemistry. 39 (51): 16016–25. doi:10.1021/bi001754t. PMID 11123929.
- Catalano A, Romano M, Robuffo I, Strizzi L, Procopio A (August 2001). "Methionine aminopeptidase-2 regulates human mesothelioma cell survival: role of Bcl-2 expression and telomerase activity". Am. J. Pathol. 159 (2): 721–31. doi:10.1016/S0002-9440(10)61743-9. PMC 1850571. PMID 11485930.
- Endo H, Takenaga K, Kanno T, et al. (2002). "Methionine aminopeptidase 2 is a new target for the metastasis-associated protein, S100A4". J. Biol. Chem. 277 (29): 26396–402. doi:10.1074/jbc.M202244200. PMID 11994292.
- Kanno T, Endo H, Takeuchi K, et al. (2002). "High expression of methionine aminopeptidase type 2 in germinal center B cells and their neoplastic counterparts". Lab. Invest. 82 (7): 893–901. doi:10.1097/01.LAB.0000020419.25365.C4. PMID 12118091.
- Datta R, Tammali R, Datta B (2003). "Negative regulation of the protection of eIF2alpha phosphorylation activity by a unique acidic domain present at the N-terminus of p67". Exp. Cell Res. 283 (2): 237–46. doi:10.1016/S0014-4827(02)00042-3. PMID 12581743.
- Serero A, Giglione C, Sardini A, Martinez-Sanz J, Meinnel T (December 2003). "An unusual peptide deformylase features in the human mitochondrial N-terminal methionine excision pathway". J. Biol. Chem. 278 (52): 52953–63. doi:10.1074/jbc.M309770200. PMID 14532271.
- Selvakumar P, Lakshmikuttyamma A, Kanthan R, Kanthan SC, Dimmock JR, Sharma RK (April 2004). "High expression of methionine aminopeptidase 2 in human colorectal adenocarcinomas". Clin. Cancer Res. 10 (8): 2771–5. doi:10.1158/1078-0432.CCR-03-0218. PMID 15102683.
- Kim S, LaMontagne K, Sabio M, Sharma S, Versace RW, Yusuff N, Phillips PE (May 2004). "Depletion of methionine aminopeptidase 2 does not alter cell response to fumagillin or bengamides". Cancer Res. 64 (9): 2984–7. doi:10.1158/0008-5472.CAN-04-0019. PMID 15126329.
PDB gallery | |
|---|---|
|