Irish elk
The Irish elk (Megaloceros giganteus)[1][2] also called the giant deer or Irish deer, is an extinct species of deer in the genus Megaloceros and is one of the largest deer that ever lived. Its range extended across Eurasia during the Pleistocene, from Ireland to Lake Baikal in Siberia. The most recent remains of the species have been carbon dated to about 7,700 years ago in Siberia.[3][4]
| Irish elk | |
|---|---|
.jpg) | |
| Mounted skeleton | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Artiodactyla |
| Family: | Cervidae |
| Subfamily: | Cervinae |
| Genus: | †Megaloceros |
| Species: | †M. giganteus |
| Binomial name | |
| †Megaloceros giganteus (Blumenbach, 1799) | |
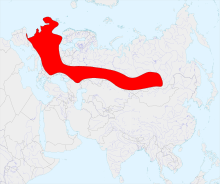 | |
| Time averaged range of M. giganteus during the Late Pleistocene | |
| Synonyms | |
| |
Although abundant skeletal remains have been found in bogs in Ireland, the animal was not exclusive to Ireland nor closely related to either of the living species currently called elk: Alces alces (the European elk, known in North America as the moose) or Cervus canadensis (the North American elk or wapiti). For this reason, the name "giant deer" is used in some publications, instead of "Irish elk".[5][6][7][8][9] Although one study suggested that the Irish elk was closely related to the red deer (Cervus elaphus),[10] most other phylogenetic analyses support that their closest living relatives are fallow deer (Dama dama).[8][11][12][13]
Taxonomy
Research history

The first scientific descriptions of the animal's remains were made by Irish physician Thomas Molyneux in 1695, who identified large antlers from Dardistown, Dublin—which were apparently commonly unearthed in Ireland—as belonging to the Elk (known as the Moose in North America), concluding that it was once abundant on the island.[14] It was first formally named as Alce gigantea by Johann Friedrich Blumenbach in his Handbuch der Naturgeschichte in 1799,[15] with Alce being a variant of Alces, the Latin name for the Elk. French scientist Georges Cuvier documented in 1812 that the Irish elk did not belong to any species of mammal currently living, declaring it "le plus célèbre de tous les ruminans fossiles".[16] In 1827 Joshua Brookes, in a listing of his zoological collection, named the new genus Megaloceros (spelled Megalocerus in the earlier editions) in the following passage:[17][18]
Amongst other Fossil Bones , there [are] ... two uncommonly fine Crania of the Megalocerus antiquorum (Mihi). (Irish), with unusually fine horns, (in part restored)
— Joshua Brookes, Brookesian Museum. The Museum of Joshua Brookes, Esq. Anatomical and Zoological Preparations, p 20.
The etymology being from Greek: μεγαλος megalos "great" + κερας keras "horn, antler".[19] The type and only species named in the description being Megaloceros antiquorum, based on Irish remains now considered to belong to M. giganteus, making the former a junior synonym. The original description was considered by Adrian Lister in 1987 to be inadequate for a taxonomic definition.[2] In 1828 Brookes published an expanded list in the form of a cataologue for an upcoming auction, which included the Latin phrase "Cornibus deciduis palmatis" as a description of the remains. The 1828 publication was approved by International Commission on Zoological Nomenclature (ICZN) in 1977 as an available publication for the basis of zoological nomenclature.[2] Adrian Lister in 1987 judged that "the phase "Cornibus deciduis palmatis" constitutes a definition sufficient under the [International Code of Zoological Nomenclature] (article 12) to validate Megalocerus."[2] The original spelling of Megalocerus was never used after its original publication.[18]

In 1844 Richard Owen named another synonym of the Irish elk, including it within the newly named subgenus Megaceros, Cervus (Megaceros) hibernicus. This has been suggested to be derived from another junior synonym of the Irish elk described by J. Hart in 1825, Cervus megaceros.[2] Despite being a junior synonym, Megaloceros remained in obscurity and Megaceros became the common genus name for the taxon.[18] The combination "Megaceros giganteus" was in use by 1871.[20] George Gaylord Simpson in 1945 revived the original Megaloceros name, which became progressively more widely used, until a taxonomic decision in 1989 by the ICZN confirmed the priority of Megaloceros over Megaceros, and Megaloceros to be the correct spelling.[18][21]
Before the 20th century, the Irish elk, having evolved from smaller ancestors with smaller antlers, was taken as a prime example of orthogenesis (directed evolution), an evolutionary mechanism opposed to Darwinian evolution in which the successive species within the lineage become increasingly modified in a single undeviating direction, evolution proceeding in a straight line void of natural selection. Orthogenesis was claimed to have caused an evolutionary trajectory towards antlers that became larger and larger, eventually causing the species' extinction because the antlers grew to sizes which inhibited proper feeding habits and caused the animal to become trapped in tree branches.[8] In the 1930s, orthogenesis was disputed by Darwinians led by Julian Huxley, who noted that antler size was not grossly large, and was proportional to body size.[22][23] The currently favoured view is that sexual selection was the driving force behind the large antlers rather than orthogenesis or natural selection.[23]

Evolution

M. giganteus belongs to the genus Megaloceros. Megaloceros is a member of the possibly polyphyletic (invalid) tribe "Megalocerini" or "Megacerini", alongside Megaceroides, Praemegaceros, Eucladoceros and Sinomegaceros, which are often collectively referred to as "giant deer". The taxonomy of giant deer is controversial, with genus names used for species varying substantially between authors.[24][25] The earliest possible record of the genus is a partial antler from the Early Pleistocene MN 17 (2.5–1.8 Ma) of Stavropol Krai in the North Causcaus of Russia, which were given the name of M. stavropolensis in 2016,[26] however these have been subsequently suggested to belong to Arvernoceros instead.[24][25] The oldest generally accepted records of the genus are from the late Early Pleistocene.[27] Other species often considered to belong to Megaloceros include the reindeer sized M. savini, which is known from early Middle Pleistocene (~700,000–450,000 years ago) localities in England, France, Spain and Germany, and the more recently described species M. novocarthaginiensis, which is known from late Early Pleistocene (0.9–0.8 Ma) localities in Spain, and the small M. matritensis endemic to the Iberian peninsula during the late Middle Pleistocene (~400,000 to 250,000 years ago), which overlaps chronologically with the earliest M. giganteus records. Jan van der Made proposed these species to be chronospecies, due to shared morphological characteristics not found in M. giganteus and gradual transition of morphological characters through time.[24] M. savini has also been suggested to comprise the separate genus Praedama by some scholars, though they are often considered closely related. Roman Croitor has suggested closer affinities to Eucladoceros.[25]
The origin of M. giganteus remains unclear, and appears to lie outside Western Europe.[24] Jan van der Made has suggested that remains of an indeterminate Megaloceros species from the late Early Pleistocene (~1.2 Ma) of Libakos in Greece are closer to M. giganteus than the M. novocarthaginiensis-matritensis lineage due to the shared molarisation of the lower fourth premolar (P4).[24] Croitor has suggested that M. giganteus is closely related to what was originally described as Dama clactoniana mugharensis (which he proposes be named M. mugharensis) from the Middle Pleistocene of Tabun Cave in Israel, due to similarities in the antlers, molars and premolars.[25] The earliest possible records of M. giganteus comes from Homersfield, Norfolk, thought to be about 450,000 years ago—though the dating is uncertain.[28] The oldest securely dated Middle Pleistocene records are those from Hoxne, which have been dated to MIS 11 (424,000 to 374,000 years ago),[29][24] other Middle Pleistocene early records include Steinheim an der Murr, Germany, (classified as M. g. antecedens) about 400,000–300,000 years ago and Swanscombe.[28][24] Most remains of the Irish elk are known from the Late Pleistocene. A large proportion of the known remains of M. giganteus are from Ireland, which mostly date to the Allerød oscillation near the end of the Late Pleistocene around 13,000 years ago. Over 100 individuals have been found in Ballybetagh Bog near Dublin.[30]
It has been historically thought that, because both have palmated antlers, the Irish elk and fallow deer (Dama spp.) are closely related, this is supported by several other morphological similarities, including the lack of upper canines, proportionally long braincase and nasal bones, and proportionally short front portion of the skull.[25] In 2005, two fragments of mitochondrial DNA (mtDNA) from the cytochrome b gene were extracted and sequenced from 4 antlers and a bone, the mtDNA found that the Irish Elk was nested within Cervus, and were inside the clade containing living red deer (Cervus elaphus). Based on this, the authors suggested that the Irish elk and red deer interbred, which would be unsurprising as hybridisation is known to occur among present-day deer species.[10] However, another study from the same year in Nature utilising both fragmentary Mitochondrial DNA and morphological data found that the Irish Elk was indeed most closely related to Dama.[11] The close relationship with Dama was supported by another cytochrome b study in 2006,[8] a 2015 study involving the full mitochondrial genome,[12] and by a 2017 morphological analysis of the bony labyrinth.[13] The 2006 and 2017 studies also directly suggest that the results of the 2005 cytochrome b paper were the result of DNA contamination.
Description


The Irish elk stood about 2.1 m (6 ft 11 in) tall at the shoulders and carried the largest antlers of any known deer, a maximum of 3.65 m (12.0 ft) from tip to tip and 40 kg (88 lb) in weight. For body size, at about 450–600 kg (990–1,300 lb) and up to 700 kg (1,500 lb) or more,[32][33][34] the Irish elk was the heaviest known cervine ("Old World deer");[11] and tied with the extant Alaska moose (Alces alces gigas) as the third largest known deer. The second and first largest are Cervalces scotti and C. latifrons, respectively.[32][33] Nonetheless, compared to Alces, Irish elk appear to have had a more robust skeleton, with older and more mature Alces skeletons bearing some resemblance to those of prime Irish elk, and younger Irish elk resembling prime Alces. Likely due to different social structures, the Irish elk exhibits more marked sexual dimorphism than Alces, with Irish elk bucks being notably larger than does.[35] In total, Irish elk bucks may have ranged from 450–700 kg (990–1,540 lb), with an average of 575 kg (1,268 lb), and does may have been relatively large, about 80% of buck size, or 460 kg (1,010 lb) on average.[36] The distingushing characters of M. giganteus include concave frontals, proportionally long braincase, proportionally short front section of the skull (orbitofrontal region), alongside the absence of upper canines and the molarisation of the lower fourth premolar (P4). The skull and mandible of the Irish elk exhibit substantial thickening (pachyostosis), with the early and complete obliteration of cranial sutures.[25]
Based on Upper Palaeolithic cave paintings, the Irish elk seems to have had overall light colouration, with a dark stripe running along the back, a stripe on either side from shoulder to haunch, a dark collar on the throat and a chinstrap, and a dark hump on the withers (between the shoulder blades). In 1989, American palaeontologist Dale Guthrie suggested that, like bison, the hump allowed a higher hinging action of the front legs to increase stride length while running. Valerius Geist suggested that the hump may have also been used to store fat. Localising fat rather than evenly distributing it may have prevented overheating while running or in rut during the summer.[36]
Habitat
Neither exclusive to Ireland nor an elk, it was named so because the most well-known and best-preserved fossil specimens have been found in lake sediments and peat bogs in Ireland. The Irish elk had a far-reaching range, extending from the Atlantic Ocean in the West to Lake Baikal in the East. They do not appear to have extended northward onto the open mammoth steppe, rather keeping to the boreal steppe-woodland environments, which consisted of scattered spruce and pine, as well as low-lying herbs and shrubs including grasses, sedges, Ephedra and Chenopodiaceae.[4]


Palaeobiology
Physiology
In 1998, Canadian biologist Valerius Geist hypothesised that the Irish elk was cursorial (adapted for running and stamina). He noted that the Irish elk physically resembled reindeer, and the body proportions of the Irish elk are similar to those of the cursorial addax, oryx, and saiga antelope. These include the relatively short legs, the long front legs nearly as long as the hind legs, and a robust and cylindrical body. Cursorial saiga, gnus, and reindeer have a top speed of over 80 km/h (50 mph), and can maintain high speeds for up to 15 minutes.[36]
Reproduction
At Ballybetagh Bog, over 100 Irish elk individuals were found, all small antlered bucks. This indicates that bucks and does segregated during at least winter and spring. Many modern deer species do this in part because males and females have different nutritional requirements and need to consume different types of plants. Segregation would also imply a polygynous society, with stags fighting for control over harems during rut. Because most of the individuals found were juvenile or geriatric and were likely suffering from malnutrition, they probably died from winterkill. It is possible that most Irish elk specimens known had died from winterkill, and winterkill is the highest source of mortality among many modern deer species. Bucks generally suffer higher mortality rates because they eat little during the autumn rut.[37] For rut, a lean stag normally 575 kg (1,268 lb) may have fattened up to 690 kg (1,520 lb), and would burn through the extra fat over the next month.[36]
Assuming a similar response to starvation as red deer, a large, healthy Irish elk stag with 40 kg (88 lb) antlers would have had 20-to-28 kg (44-to-62 lb) antlers under poor conditions;[16][33] and an average sized Irish elk stag with 35 kg (77 lb) antlers would have had 18 to 25 kg (40 to 55 lb) antlers under poorer conditions,[38] similar sizes to the moose. A similar change in a typical Irish elk population with prime stags having 35 kg (77 lb) antlers would result in antler weights of 13 kg (29 lb) or less in worsening climatic conditions. This is within the range of present day wapiti/red deer (Cervus spp.) antler weights.[36] Irish elk antlers vary widely in form depending upon the habitat, such as a compact, upright shape in closed forest environments.[38] Irish elk likely shed their antlers and re-grew a new pair during mating season. Antlers generally require high amounts of calcium and phosphate, especially those for stags which have larger structures, and the massive antlers of Irish elk may have required much greater quantities. Stags typically meet these requirements in part from their bones, suffering from a condition similar to osteoperosis while the antlers are growing, and replenishing them from food plants after the antlers have grown in or reclaiming nutrients from shed antlers.[33]
The large antlers have generally been explained as being used for male-male battle during mating season.[39] They may have also been used for display,[16] to attract females and assert dominance against rival males.[37] A finite element analysis of the antlers suggested that during fighting, the antlers were likely to interlock around the middle tine, the high stress when interlocking on the distal tine suggests that the fighting was likely more constrained and predictable than among extant deer, likely involving twisting motions, as is known in extant deer with palmated antlers.[40]
In deer, gestation time generally increases with body size. A 460 kg (1,010 lb) doe may have had a gestation period of about 274 days. Based on this and patterns seen in modern deer, last year's antlers in Irish elk bucks were potentially shed in early March, peak antler growth in early June, completion by mid-July, shedding velvet (a layer of blood vessels on the antlers in-use while growing them) by late July, and the height of rut falling on the second week of August. Geist, believing the Irish elk to have been a cursorial animal, concluded that a doe would have to have produced nutrient-rich milk so that her calf would have enough energy and stamina to keep up with the herd.[36]
Life history
The mesodont (meaning neither high (hypsodont) or low (brachydont) crowned) condition of the teeth suggests that the species was a mixed feeder, being able to both browse and graze. Pollen remains from teeth found in the North Sea around 43 kya were found to be dominated by Artemisia and other Asteraceae, with minor Plantago, Helianthemum, Plumbaginaceae and Salix.[41] A stable isotope analysis of the terminal Pleistocene Irish population suggests a grass and forb based diet, supplemented by browsing during stressed periods.[42] Dental wear patterns of specimens from the late Middle and Late Pleistocene of Britain suggest a diet tending towards mixed feeding and grazing, but with a wide range including leaf browsing.[43]
Based on the dietary requirements of red deer, a 675 kg (1,488 lb) lean Irish elk stag would have needed to consume 39.7 kg (88 lb) of fresh forage daily. Assuming antler growth occurred over a span of 120 days, a stag would have required 1,372 g of protein daily, as well as access to nutrient- and mineral-dense forage starting about a month before antlers began sprouting and continuing until they had fully grown. Such forage is not very common, and stags perhaps sought after aquatic plants in lakes. After antler growing, stags could probably satisfy their nutritional requirements in productive sedge lands bordered by willow and birch forests.[36]
The Irish elk may have been preyed upon by the large carnivores of the time, including the cave lion,[44] and the cave hyena.[45]
Extinction

Lister and Stewart note that "With the exception of the Irish late-glacial, finds of M. giganteus, throughout its Pleistocene history, are generally low in abundance at sites where they occur, suggesting that it was generally a rare species."[4]
Historically, the extinction of the elk has been attributed to the encumbering size of the antlers, a "maladaptation" making fleeing through forests especially difficult for males while being chased by human hunters,[16] or being too taxing nutritionally when the vegetation makeup shifted.[33] In these scenarios, sexual selection by does for stags with large antlers would have contributed to decline.[46]
However, antler size decreases through the Late Pleistocene and into the Holocene, and so may not have been the primary cause of extinction.[38] A reduction in forest density in the Late Pleistocene and a lack of sufficient high-quality forage is associated with a decrease in body and antler size.[47] Such resource constriction may have cut female fertility rates in half.[38] Human hunting may have forced Irish elk into suboptimal feeding grounds.[3]
The range of the taxon appears to have collapsed during the Last Glacial Maximum, with few remains known between 27.5 and 14.6 kya, and none between 23.3 and 17.5 kya. The remains of the taxon substantially increase during the latest Pleistocene, where it appears to have re-colonised most of its former range, with abundant remains in the UK, Ireland and Germany.[4]
While the range of the taxon was dramatically reduced after the Pleistocene-Holocene transition, it managed to survive into the early Northgrippian in the eastern part of its range within European Russia and Siberia, in a belt extending from Maloarchangelsk in the East to Preobrazhenka in the west. It is suggested that extinction was contributed to by further post Holocene transition climatic changes transforming preferred open habitat into uninhabitable dense forest.[4] The final demise may have been caused by a number of factors both on a continental and regional scale, including climate change and hunting.[47][48]
Lister and Stewart concluded in a study of the extinction of the Irish elk that "it seems clear that environmental factors, cumulatively over thousands of years, reduced giant deer populations to a highly vulnerable state. In this situation, even relatively low-level hunting by small human populations could have contributed to its extinction."[4]
Cultural significance

A handful of Irish elk depictions are known from the art of the Upper Paleolithic in Europe, but these are much less abundant than the common red deer and reindeer depictions. The bones of the Irish elk are uncommon in localities were they are found, and only a handful of examples of human interaction are known.[4] A mandible from Ofatinţi, Moldova dating to either the Eemian or the early Late Pleistocene, "is peculiar because it has ancient tool-made notches on its lateral side".[49] Several M. giganteus bones from the Chatelperronian levels of the Labeko Koba site in Spain are noted for bearing puncture marks, which have been interpreted as anthropogenic.[50] A terminal Pleistocene (13710-13215 cal BP) skull from Lüdersdorf, Germany is noted to have had the antler and facial part of the skull removed in a way unlikely to be due to natural causes.[51] A calcaneum from an associated lower hind limb from the early Holocene site of Sosnovy Tushamsky in Siberia is noted to have "two short and deep traces of cutting blows", which are interpreted as "clear evidence of butchery".[52][4] The use of shed antler bases is also known, at the terminal Pleistocene (Allerød) Endingen VI site in Germany, a shed antler base appears to have been used in a way analogous to a lithic core to produce "blanks" for the manufacture of barbed projectile tips.[53][4] A ring like mark on a shed antler beam from the similarly aged Paderborn site in Germany has been suggested to be anthropogenic.[54]
Due to the abundance of Irish elk remains in Ireland, a thriving trade in their bones existed there during the 19th century to supply museums and collectors. Skeletons and skulls with attached antlers were also prized ornaments in aristocratic homes. The remains of Irish elk were of high value: "In 1865, full skeletons might fetch £30, while particularly good heads with antlers could cost £15." with £15 being more than 30 weeks wages for a low skilled worker at the time.[55]
A folk memory of the Irish elk was once thought to be preserved in the Middle High German word Schelch, a large beast mentioned in the 13th-century Nibelungenlied along with the then-extant aurochs (Dar nach schluch er schiere, einen Wisent und einen Elch, Starcher Ure vier, und einen grimmen Schelch / "After this he straightway slew a Bison and an Elk, Of the strong Wild Oxen four, and a single fierce Schelch.") This opinion is no longer widely held.[16] The Middle Irish word segh was also suggested as a reference to the Irish elk.[56][57] Turf cutters of Clooney and Tulla in County Clare, Ireland referred to the Irish elk as the Fiaghmore.[58] However, these interpretations are not conclusive.[59]
See also

References
- Geist, Valerius (1998). "Megaloceros: The Ice Age Giant and Its Living Relatives". Deer of the World: Their Evolution, Behaviour, and Ecology. Stackpole Books. ISBN 978-0-8117-0496-0.
- Lister, A.M. (1987). "Megaceros or Megaloceros? The nomenclature of the giant deer". Quaternary Newsletter. 52: 14–16.
- Stuart, A.J.; Kosintsev, P.A.; Higham, T.F.G.; Lister, A.M. (2004). "Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth" (PDF). Nature. 431 (7009): 684–689. Bibcode:2004Natur.431..684S. doi:10.1038/nature02890. PMID 15470427. Archived from the original (PDF) on 14 September 2006. Supplementary information. Erratum in Stuart, A. J. (2005). "Erratum: Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth". Nature. 434 (7031): 413. Bibcode:2005Natur.434..413S. doi:10.1038/nature03413.
- Lister, Adrian M.; Stuart, Anthony J. (January 2019). "The extinction of the giant deer Megaloceros giganteus (Blumenbach): New radiocarbon evidence". Quaternary International. 500: 185–203. Bibcode:2019QuInt.500..185L. doi:10.1016/j.quaint.2019.03.025.
- Vislobokova, I. A. (2010), "Giant Deer: Origin, Evolution, Role in the Biosphere", Paleontological Journal. vol. 46 no. 7 pg. 643–775
- Bro-Jorgensen, J. (2014), "Will their armaments be their downfall? Large horn size increases extinction risk in bovids," Animal Conservation. vol. 17 no. 1 pg. 80–87
- Lemaitre, J. F. (2014), "The allometry between secondary sexual traits and body size is nonlinear among cervids", Biology Letter. vol. 10 no. 3
- Hughes, Sandrine; Hayden, Thomas J.; Douady, Christophe J.; Tougard, Christelle; Germonpré, Mietje; Stuart, Anthony; Lbova, Lyudmila; Carden, Ruth F.; Hänni, Catherine; Say, Ludovic (2006). "Molecular phylogeny of the extinct giant deer, Megaloceros giganteus". Molecular Phylogenetics and Evolution. 40 (1): 285–291. doi:10.1016/j.ympev.2006.02.004. PMID 16556506.
- Vislobokova, I. A. (2011), "Historical Development and Geographical Distribution of Giant Deer (Cervidae, Megacerini)", Paleontological Journal. vol. 45 no. 6 pg. 674–688
- Kuehn, Ralph; Ludt, Christian J.; Schroeder, Wolfgang; Rottmann, Oswald (2005). "Molecular Phylogeny of Megaloceros giganteus — the Giant Deer or Just a Giant Red Deer?". Zoological Science. 22 (9): 1031–1044. doi:10.2108/zsj.22.1031. PMID 16219984.
- Lister, Adrian M.; Edwards, Ceiridwen J.; Nock, D. A. W.; Bunce, Michael; van Pijlen, Iris A.; Bradley, Daniel G.; Thomas, Mark G.; Barnes, Ian (2005). "The phylogenetic position of the giant deer Megaloceros giganteus". Nature. 438 (7069): 850–853. Bibcode:2005Natur.438..850L. doi:10.1038/nature04134. PMID 16148942.
- Immel, Alexander; Drucker, Dorothée G.; Bonazzi, Marion; Jahnke, Tina K.; Münzel, Susanne C.; Schuenemann, Verena J.; Herbig, Alexander; Kind, Claus-Joachim; Krause, Johannes (2015). "Mitochondrial Genomes of Giant Deers Suggest their Late Survival in Central Europe". Scientific Reports. 5 (10853): 10853. Bibcode:2015NatSR...510853I. doi:10.1038/srep10853. PMC 4459102. PMID 26052672.
- Mennecart, Bastien; DeMiguel, Daniel; Bibi, Faysal; Rössner, Gertrud E.; Métais, Grégoire; Neenan, James M.; Wang, Shiqi; Schulz, Georg; Müller, Bert; Costeur, Loïc (13 October 2017). "Bony labyrinth morphology clarifies the origin and evolution of deer". Scientific Reports. 7 (1): 13176. Bibcode:2017NatSR...713176M. doi:10.1038/s41598-017-12848-9. ISSN 2045-2322. PMC 5640792. PMID 29030580.
- Molyneux, T. (1695). "A Discourse Concerning the Large Horns Frequently Found under Ground in Ireland, Concluding from Them That the Great American Deer, Call'd a Moose, Was Formerly Common in That Island: With Remarks on Some Other Things Natural to That Country". Philosophical Transactions of the Royal Society. 19 (227): 489–512. Bibcode:1695RSPT...19..489M. doi:10.1098/rstl.1695.0083.
- Blumenbach J. 1799. Handbuch der Naturgeschichte (6th Ed.) 16: 697
- Gould, Stephen Jay (1974). "The Origin and Function of 'Bizarre' Structures: Antler Size and Skull Size in the 'Irish Elk,' Megaloceros giganteus". Evolution. 28 (2): 191–220. doi:10.2307/2407322. JSTOR 2407322. PMID 28563271.
- Joshua Brookes (1827) "Brookesian Museum. The Museum of Joshua Brookes, Esq. Anatomical and Zoological Preparations" London Gold and Walton
- Lister, A M, 1987 Megaloceros Brookes 1828 Mammalia Artiodactyla Proposed Emendation Of The Original Spelling The Bulletin of zoological nomenclature. 44 255–256
- "Megaloceros". Oxford Dictionary. Retrieved 12 July 2020 – via Lexico.
- Vogt, Carl (1871). Lehrbuch der Geologie und Petrefactenkunde : zum Gebrauche bei Vorlesungen und zum Selbstunterrichte. p. 7. OCLC 162473843.
- International Commission on Zoological Nomenclature 1989. Opinion 1566. Megaloceros Brookes, 1828 (Mammalia, Artiodactyla): original spelling emended. Bulletin of zoological nomenclature 46: 219–220.
- Anderson, Kristina. "What On Earth - A Canadian Newsletter for the Earth Sciences". What On Earth - A Canadian Newsletter for the Earth Sciences. N.p., 15 November 2002. Web. 23 October 2014.
- Zimmer, Carl. "The Allure of Big Antlers". The Loom. Discover, National Geographic. 3 September 2008. Web. 23 October 2014.
- Van der Made, Jan. 2019. The dwarfed "giant deer" Megaloceros matritensis n.sp. from the Middle Pleistocene of Madrid - A descendant of M. savini and contemporary to M. giganteus. Quaternary International 520. 110–139. Accessed 2019-02-04.
- Croitor, Roman (2018). Plio-Pleistocene deer of Western Palearctic : taxonomy, systematics, phylogeny. pp. 72 (stavropolensis) 93-94 (Praedama) 100-101 (Megaloceros) 105 (mugharensis). ISBN 978-9975-66-609-1. OCLC 1057238213.
- Titov, V. V.; Shvyreva, A. K. (January 2016). "Deer of the genus Megaloceros (Mammalia, Cervidae) from the Early Pleistocene of Ciscaucasia". Paleontological Journal. 50 (1): 87–95. doi:10.1134/S0031030116010111. ISSN 0031-0301.
- van der Made, J.; Tong, H.W. (March 2008). "Phylogeny of the giant deer with palmate brow tines Megaloceros from west and Sinomegaceros from east Eurasia". Quaternary International. 179 (1): 135–162. Bibcode:2008QuInt.179..135V. doi:10.1016/j.quaint.2007.08.017.
- Lister, Adrian M. (September 1994). "The evolution of the giant deer, Megaloceros giganteus (Blumenbach)". Zoological Journal of the Linnean Society. 112 (1–2): 65–100. doi:10.1111/j.1096-3642.1994.tb00312.x.
- Ashton, Nick; Lewis, Simon G.; Parfitt, Simon A.; Penkman, Kirsty E.H.; Russell Coope, G. (April 2008). "New evidence for complex climate change in MIS 11 from Hoxne, Suffolk, UK". Quaternary Science Reviews. 27 (7–8): 652–668. doi:10.1016/j.quascirev.2008.01.003. PMC 2748712. PMID 19777138.
- Johnston, Penny; Kelly, Bernice; Tierney, John. "Megaloceros Giganteus on the Loose" (PDF). Seanda: The NRA Archaeology Magazine. pp. 58–59. Archived from the original (PDF) on 29 October 2013.
- Jaubert, Jacques. L'" art " pariétal gravettien en France : éléments pour un bilan chronologique. p. 446. OCLC 803593335.
- R. D. E. Mc Phee, Extinctions in Near Time: Causes, Contexts, and Consequences p. 262
- Moen, Ron A.; Pastor, John; Cohen, Yosef (1999). "Antler growth and extinction of Irish elk" (PDF). Evolutionary Ecology Research: 235–249.
- A View to a Kill: Investigating Middle Palaeolithic Subsistence Using an Optimal Foraging Perspective
- Breda, M. (2005). "The morphological distinction between the postcranial skeleton of Cervalces/Alces and Megaloceros giganteus and comparison between the two Alceini genera from the Upper Pliocene–Holocene of Western Europe". Geobios. 38 (2): 151–170. doi:10.1016/j.geobios.2003.09.008.
- Geist, V. (1998). "Megaloceros: the ice age giant and its living relatives". Deer of the World: Their Evolution, Behaviour, and Ecology. Stackpole Books. ISBN 978-0-8117-0496-0.
- Barnosky, Anthony D. (19 April 1985). "Taphonomy and Herd Structure of the Extinct Irish Elk, Megalocerous giganteus". Science. New. 228 (4697): 340–344. Bibcode:1985Sci...228..340B. doi:10.1126/science.228.4697.340. PMID 17790237.
- O'Driscoll Worman, Cedric; Kimbrell, Tristan (2008). "Getting to the hart of the matter: Did antlers truly cause the extinction of the Irish elk?". Oikos. 117 (9): 1397–1405. doi:10.1111/j.0030-1299.2008.16608.x.
- Kitchener, A. (1987). "Fighting behavior of the extinct Irish elk". Modern Geology. 11: 1–28.
- Klinkhamer, Ada J.; Woodley, Nicholas; Neenan, James M.; Parr, William C. H.; Clausen, Philip; Sánchez-Villagra, Marcelo R.; Sansalone, Gabriele; Lister, Adrian M.; Wroe, Stephen (9 October 2019). "Head to head: the case for fighting behaviour in Megaloceros giganteus using finite-element analysis". Proceedings of the Royal Society B: Biological Sciences. 286 (1912): 20191873. doi:10.1098/rspb.2019.1873. ISSN 0962-8452. PMC 6790765. PMID 31594504.
- van Geel, B.; Sevink, J.; Mol, D.; Langeveld, B. W.; van der Ham, R. W. J. M.; van der Kraan, C. J. M.; van der Plicht, J.; Haile, J. S.; Rey-Iglesia, A.; Lorenzen, E. D. (November 2018). "Giant deer ( Megaloceros giganteus ) diet from Mid-Weichselian deposits under the present North Sea inferred from molar-embedded botanical remains: Giant Deer Diet from Mid-Weichselian Deposits". Journal of Quaternary Science. 33 (8): 924–933. doi:10.1002/jqs.3069.
- Chritz, Kendra L.; Dyke, Gareth J.; Zazzo, Antoine; Lister, Adrian M.; Monaghan, Nigel T.; Sigwart, Julia D. (November 2009). "Palaeobiology of an extinct Ice Age mammal: Stable isotope and cementum analysis of giant deer teeth". Palaeogeography, Palaeoclimatology, Palaeoecology. 282 (1–4): 133–144. Bibcode:2009PPP...282..133C. doi:10.1016/j.palaeo.2009.08.018.
- Rivals, Florent; Lister, Adrian M. (August 2016). "Dietary flexibility and niche partitioning of large herbivores through the Pleistocene of Britain". Quaternary Science Reviews. 146: 126. Bibcode:2016QSRv..146..116R. doi:10.1016/j.quascirev.2016.06.007.
- Stuart, A. J.; Lister, A. M. (2011). "Extinction chronology of the cave lion Panthera spelaea". Quaternary Science Reviews. 30 (17–18): 2337. Bibcode:2011QSRv...30.2329S. doi:10.1016/j.quascirev.2010.04.023.
- Diedrich, C. G. (2011). "An Ice Age spotted hyena Crocuta crocuta spelaea (Goldfuss 1823) population, their excrements and prey from the Late Pleistocene hyena den of the Sloup Cave in the Moravian Karst, Czech Republic". Historical Biology. 24 (2): 161–185. doi:10.1080/08912963.2011.591491.
- Kokko, H.; Brooks, R. (2003). "Sexy to die for? sexual selection and the risk of extinction". Annales Zoologici Fennici. 40 (2): 207–219. ProQuest 18804232.
- Gonzalez, Silvia; Andrew Kitchener; Adrian Lister (15 June 2000). "Survival of the Irish elk into the Holocene". Nature. 405 (6788): 753–754. Bibcode:2000Natur.405..753G. doi:10.1038/35015668. PMID 10866185.
- "Irish Elk Survived after Ice Age Ended" Author(s): S. P. Science News, Vol. 166, No. 19 (6 November 2004), p. 301. Society for Science & the Public.
- Croitor, Roman; Stefaniak, Krzysztof; Pawłowska, Kamilla; Ridush, Bogdan; Wojtal, Piotr; Stach, Małgorzata (April 2014). "Giant deer Megaloceros giganteus Blumenbach, 1799 (Cervidae, Mammalia) from Palaeolithic of Eastern Europe". Quaternary International. 326–327: 91–104. doi:10.1016/j.quaint.2013.10.068.
- A. Villaluenga, A. Arrizabalaga, J. Rios-Garaizar "Multidisciplinary approach to two Chatelperronian series: lower IX layer of Labeko Koba and X level of Ekain (Basque Country, Spain)" Journal of Taphonomy, 10 (2012), pp. 499–520
- B. Bratlund "Ein Riesenhirschschädel mit Bearbeitungsspuren aus Lüdersdorf", Kreis Grevesmühlen. Offa 49/50 (1992/1993) (1994), pp. 7–14
- S. K. Vasiliev, V. S. Slavinsky, A. V. Postnov. "The Irish elk (Megaloceros giganteus Blumenbach, 1803) in the paleofauna of the Holocene sites of the northern Angara region (Ust-Tushama-1, Sosnovy Tushamsky Ostrov, Ust-Talaya)" Journal of Novosibirsk State University, Series: Hist. Philol., 12/7 (2013), pp. 177–185 Archaeology and Ethography
- K. Kaiser, P. De Klerk, T. Terberger. "Die 'Riesenhirsch Fundstelle' von Endingen: geowissenschaftliche und archäologische Untersuchungen an einem spätglazialen Fundplatz in Vorpommern". Eiszeitalt und Gegenwart. 49 (1999). pp. 102–123.
- M. Baales, S. Birker, H.-O. Pollman, W. Rosendahl, B. Stapel "Erstmals datierte organische Artefakte aus dem Spätpaläolithikum Westfalens Archäologie in Westfalen". Lippe, 2012 (2012), pp. 24–27
- Adelman, Juliana (April 2012). "An insight into commercial natural history: Richard Glennon, William Hinchy and the nineteenth-century trade in giant Irish deer remains". Archives of Natural History. 39 (1): 16–26. doi:10.3366/anh.2012.0059. ISSN 0260-9541.
- Cuvier, Georges; Edward Griffith; et al. (1827). The Animal Kingdom Arranged in Conformity with Its Organization. London: G. B. Whittaker. pp. 87–89.
- Hibbert, S. (October 1830 – April 1830). "Additional Contributions towards the History of the Cervus Euryce, or Fossil Elk of Ireland". The Edinburgh Journal of Science. 2 (3): 314.
- Westropp, Thomas Johnson (1913). "A Folklore Survey of County Clare. Chapter 17: Animal and Plant Superstitions".
- Owen, Richard (1846). A History of British Fossil Mammals, and Birds. London: John Van Voorst. pp. 461–462.
Further reading
- Kurten, Bjorn (1995): Dance of the Tiger. University of California Press. ISBN 0-520-20277-5.
- Kurten is a paleo-anthropologist, and in this novel he presents a theory of Neanderthal extinction. Irish elk feature prominently, under the name shelk which Kurten coins (based on the aforementioned old German schelch) to avoid the problematic aspects of "Irish" and "elk" as discussed above. The book was first published in 1980, when the name "Giant Deer" was not yet being used widely.
- Zoological Science 22: 1031–1044 (2005).
- Larson, Edward J. (2004). Evolution: The Remarkable History of a Scientific Theory.
External links
| Wikimedia Commons has media related to Megaloceros giganteus. |
- "Extinct Giant Deer Survived Ice Age, Study Says". National Geographic.
- "CGI picture from Walking with Beasts". Discovery Channel. Archived from the original on 17 February 2006. Retrieved 14 February 2006.
- "Megaloceros, Irish elk, Giant deer". BBC. Retrieved 25 October 2005.
- "The Case of the Irish Elk". University of California at Berkeley. Archived from the original on 11 November 2005. Retrieved 25 October 2005.