Mammoth steppe
During the Last Glacial Maximum, the mammoth steppe was the Earth’s most extensive biome. It spanned from Spain eastward across Eurasia to Canada and from the arctic islands southward to China.[2][3][4][5][6] It had a cold, dry climate;[7][6] the vegetation was dominated by palatable high-productivity grasses, herbs and willow shrubs,[3][6][8] and the animal biomass was dominated by bison, horses, and woolly mammoth.[7] This ecosystem covered wide areas of the northern part of the globe, thrived for approximately 100,000 years without major changes, and then suddenly became all but extinct about 12,000 years ago.[7]

Naming
At the end of the 19th century, Alfred Nehring (1890)[9] and Jan Czerski (Iwan Dementjewitsch Chersky, 1891)[10] proposed that during the last glacial period a major part of northern Europe had been populated by large herbivores and that a steppe climate had prevailed there.[11] In 1982, the scientist R. Dale Guthrie coined the term "mammoth steppe" for this paleoregion.[12][11]
Origin of the mammoth steppe
The last glacial period, commonly referred to as the 'Ice Age', spanned from 126,000 YBP–11,700 YBP[13] and was the most recent glacial period within the current ice age which occurred during the last years of the Pleistocene epoch.[14] This arctic environment was very cold and dry and probably dusty, resembling mountaintop environments, and was very different from today's swampy tundra.[15] It reached its peak during the last glacial maximum, when ice sheets commenced advancing from 33,000 years BP and reached their maximum positions 26,500 years BP. Deglaciation commenced in the Northern Hemisphere approximately 19,000 years BP, and in Antarctica approximately 14,500 years BP, which is consistent with evidence that it was the primary source for an abrupt rise in the sea level at that time.[16]
During the peak of the last glacial maximum, a vast mammoth steppe stretched from the Iberian Peninsula across Eurasia and over the Bering land bridge into Alaska and the Yukon where it was stopped by the Wisconsin glaciation. This land bridge existed because more of the planet's water was locked up in ice than now and therefore the sea levels were lower. When the sea levels began to rise this bridge was inundated around 11,000 years BP.[17]
During glacial periods, there is clear evidence for intense aridity due to water being held in glaciers and their associated effects on climate.[18][19][7] The mammoth steppe was like a huge 'inner court' that was surrounded on all sides by moisture-blocking features: massive continental glaciers, high mountains, and frozen seas. These kept rainfall low and created more days with clear skies than are seen today, which increased evaporation in the summer leading to aridity, and radiation of warmth from the ground into the black night sky in the winter leading to cold.[7] This is thought to have been caused by seven factors:
- The driving force for the core Asian steppe was an enormous and stable high-pressure system north of the Tibetan Plateau.
- Deflection of the larger portion of the Gulf Stream southward, past southern Spain onto the coast of Africa, reduced temperatures (hence moisture and cloud cover) that the North Atlantic Current brings to Western Europe.
- Growth of the Scandinavian ice sheet created a barrier to North Atlantic moisture.
- Icing over of the North Atlantic sea surface with reduced flow of moisture from the east.
- The winter (January) storm track seems to have swept across Eurasia on this axis.
- Lowered sea levels exposed a large continental shelf to the north and east producing a vast northern plain which increased the size of the continent to the north.
- North American glaciers shielded interior Alaska and the Yukon Territory from moisture flow. These physical barriers to moisture flow created a vast arid basin or protected 'inner court' spanning parts of three continents.[7]
Environment (or Biota)
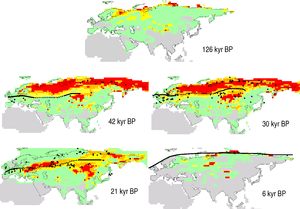
Animal biomass and plant productivity of the mammoth steppe were similar to today's African savannah.[6] There is no comparison to it today.[7][6]
Plants
The paleo-environment changed across time,[21] a proposal that is supported from mammoth dung samples found in northern Yakutia.[22] During Pleniglacial interstadials, alder, birch, and pine trees survived in northern Siberia, however during the Last Glacial Maximum only a treeless steppe vegetation existed. At the onset of the Late Glacial Interstadial (15,000–11,000 BP), global warming resulted in shrub and dwarf birch in northeastern Siberia, which was then colonized by open woodland with birch and spruce during the Younger Dryas (12,900–11,700 YBP). By the Holocene (10,000 YBP), patches of closed larch and pine forests developed.[22] Past researchers had once assumed that the mammoth steppe was very unproductive because they had assumed that its soils had a very low carbon content; however, these soils (yedoma) were preserved in the permafrost of Siberia and Alaska and are the largest reservoir of organic carbon known. It was a highly productive environment.[6][23] The vegetation was dominated by palatable high-productivity grasses, herbs and willow shrubs.[3][6][8] Herbs were far more widespread than they are today, and were the main food source of the large plant eating mammals.[24]








Animals
The mammoth steppe was dominated in biomass by bison, horse, and the woolly mammoth, and was the center for the evolution of the Pleistocene woolly fauna.[7] Notable carnivores found across the whole range of the mammoth steppe included Panthera spelaea, the wolf Canis lupus and the brown bear Ursus arctos. While the cave hyena was part of mammoth steppe faunas in Europe, it did not extend into the core high latitude north asian range of the biome.[26] On Wrangel Island, the remains of woolly mammoth, woolly rhinoceros, horse, bison and musk ox have been found. Reindeer and small animal remains do not preserve, but reindeer excrement has been found in sediment.[6] In the most arid regions of the mammoth steppe that were to the south of Central Siberia and Mongolia, woolly rhinoceros were common[27][6] but woolly mammoths were rare.[28][6] Reindeer live in the far north of Mongolia today and historically their southern boundary passed through Germany and along the steppes of eastern Europe,[29][6] indicating they once covered much of the mammoth steppe.[6] Mammoths survived on the Taimyr Peninsula until the Holocene.[8][6] A small population of mammoths survived on St. Paul Island, Alaska, up until 3750 BC,[30][31] and the small[32] mammoths of Wrangel Island survived until 1650 BC.[33][34] Bison in Alaska and the Yukon, and horses and muskox in northern Siberia, have survived the loss of the mammoth steppe.[6] One study has proposed that a change of suitable climate caused a significant drop in the mammoth population size, which made them vulnerable to hunting from expanding human populations. The coincidence of both of these impacts in the Holocene most likely set the place and time for the extinction of the woolly mammoth.[20]
Decline of the mammoth steppe
The mammoth steppe had a cold, dry climate.[7][6] During the past interglacial warmings, forests of trees and shrubs expanded northward into the mammoth steppe, when northern Siberia, Alaska and the Yukon (Beringia) would have formed a mammoth steppe refugium. When the planet grew colder again, the mammoth steppe expanded.[6] This ecosystem covered wide areas of the northern part of the globe, thrived for approximately 100,000 years without major changes, and then suddenly became extinct about 12,000 years ago.[7]
There are two theories about the decline of the mammoth steppe.
Climate change
The Climatic Hypothesis assumes that the vast mammoth ecosystem could have only existed within a certain range of climatic parameters. At the beginning of the Holocene 10,000 years ago, mossy forests, tundra, lakes and wetlands displaced mammoth steppe. It has been assumed that in contrast to other previous interglacials the cold dry climate switched to a warmer wetter climate that, in turn, caused the disappearance of the grasslands and their dependent megafauna.[3]
The extinct steppe bison (Bison priscus) survived across the northern region of central eastern Siberia until 8000 years ago. A study of the frozen mummy of a steppe bison found in northern Yakutia, Russia indicated that it was a pasture grazer in a habitat that was becoming dominated by shrub and tundra vegetation. Higher temperature and rainfall led to a decrease in its previous habitat during the early Holocene, and this led to population fragmentation followed by extinction.[35]
In 2017 a study looked at the environmental conditions across Europe, Siberia and the Americas from 25,000–10,000 YBP. The study found that prolonged warming events leading to deglaciation and maximum rainfall occurred just prior to the transformation of the rangelands that supported megaherbivores into widespread wetlands that supported herbivore-resistant plants. The study proposes that moisture-driven environmental change led to the megafaunal extinctions, and that Africa's trans-equatorial position allowed rangeland to continue to exist between the deserts and the central forests; therefore fewer megafauna species became extinct there.[36]
Human predation
The Ecosystem Hypothesis assumes that the vast mammoth ecosystem extended over a range of many regional climates and was not affected by climate fluctuations. Its highly productive grasslands were maintained by animals trampling any mosses and shrubs, and actively transpiring grasses and herbs dominated. At the beginning of the Holocene the rise in precipitation was accompanied by increased temperature, and so its climatic aridity did not change substantially. As a result of human hunting, the decreasing density of the animals was not enough to maintain the grasslands, leading to an increase in forests, shrubs and mosses with further animal reduction due to loss of feed. The mammoth continued to exist on isolated Wrangel Island until a few thousand years ago, and some of the other megafauna from that time still exist today, which indicates that something other than climate change was responsible for megafaunal extinctions.[6]
Remains of mammoth that had been hunted by humans 45,000 YBP have been found at Yenisei Bay in the central Siberian Arctic.[37] Two other sites in the Maksunuokha River valley to the south of the Shirokostan Peninsula, northeast Siberia, dated between 14,900 and 13,600 years ago showed the remains of mammoth hunting and the production of micro-blades similar to those found in northwest North America, suggesting a cultural connection.[38]
Last remnants

During the Holocene, the arid-adapted species became extinct or were reduced to minor habitats.[7] Cold and dry conditions similar to the last glacial period are found today in the eastern Altai-Sayan mountains of Central Eurasia,[39][1] with no significant changes occurring between the cold phase of the Pleistocene and the Holocene.[40][1] Recent paleo-biome reconstruction[41][42][1] and pollen analysis[43][44][45][1] suggest that some present-day Altai-Sayan areas could be considered the closest analogy to the mammoth steppe environment.[1] The environment of this region is considered to have been stable for the past 40,000 years. The Eastern part of the Altai-Sayan region forms a Last Glacial refugium. In both the Last Glacial and modern times, the eastern Altai-Sayan region has supported large herbivore and predator species adapted to the steppe, desert and alpine biomes where these biomes have not been separated by forest belts. None of the surviving Pleistocene mammals live in temperate forest, taiga, or tundra biomes. The areas of Ukok-Sailiugem in the southern Altai Republic, and Khar Us Nuur and Uvs Nuur (Ubsunur Hollow) in western Mongolia, have supported reindeer and saiga antelope since the glacial period.[1]
See also
- Pleistocene megafauna
- Pleistocene Park – a project to restore a small part of what once was the mammoth steppe.
- Quaternary extinction event
References
- Pavelková Řičánková, Věra; Robovský, Jan; Riegert, Jan (2014). "Ecological Structure of Recent and Last Glacial Mammalian Faunas in Northern Eurasia: The Case of Altai-Sayan Refugium". PLoS ONE. 9 (1): e85056. Bibcode:2014PLoSO...985056P. doi:10.1371/journal.pone.0085056. PMC 3890305. PMID 24454791.
- Adams, J. M.; Faure, H.; Faure-Denard, L.; McGlade, J. M.; Woodward, F. I. (1990). "Increases in terrestrial carbon storage from the Last Glacial Maximum to the Present". Nature. 348 (6303): 711–714. Bibcode:1990Natur.348..711A. doi:10.1038/348711a0.
- Guthrie, R.D. (1990). Frozen Fauna of the Mammoth Steppe. The University of Chicago Press, Chicago. ISBN 9780226159713.
- Sher, A.V., 1997. Nature restructuring in the East-Siberian Arctic at the Pleistocene Holocene boundary and its role in mammal extinction and emerging of modern ecosystems. Earth Cryosphere 1 (3e11), 21e29.
- Álvarez-Lao, Diego J.; García, Nuria (2011). "Geographical distribution of Pleistocene cold-adapted large mammal faunas in the Iberian Peninsula". Quaternary International. 233 (2): 159–170. Bibcode:2011QuInt.233..159A. doi:10.1016/j.quaint.2010.04.017.
- Zimov, S.A.; Zimov, N.S.; Tikhonov, A.N.; Chapin, F.S. (2012). "Mammoth steppe: A high-productivity phenomenon". Quaternary Science Reviews. 57: 26–45. Bibcode:2012QSRv...57...26Z. doi:10.1016/j.quascirev.2012.10.005.
- Guthrie, R.D., Origin and causes of the mammoth steppe: a story of cloud cover, woolly mammal tooth pits, buckles, and inside-out Beringia, Quaternary Science Reviews 20 (2001) 549-574
- Sher, A.V.; Kuzmina, S.A.; Kuznetsova, T.V.; Sulerzhitsky, L.D. (2005). "New insights into the Weichselian environment and climate of the East Siberian Arctic, derived from fossil insects, plants, and mammals" (PDF). Quaternary Science Reviews. 24 (5–6): 533–569. Bibcode:2005QSRv...24..533S. doi:10.1016/j.quascirev.2004.09.007.
- Nehring, A.: Über Tundren und Steppen der Jetzt- und Vorzeit: mit besonderer Berücksichtigung ihrer Fauna. In: F. Dümmler, Berlin 1890 .
- Chersky, I.D. (1891). "Description of the post-Tertiary mammal collection found byNew Siberian expedition 1885–1886". Notes of Russian Academy of Science. 65: 706.
- Blinnikov, Mikhail S.; Gaglioti, Benjamin; Walker, Donald A.; Wooller, Matthew J.; Zazula, Grant D. (2011). "Pleistocene graminoid-dominated ecosystems in the Arctic". Quaternary Science Reviews. 30: 2906–2929. doi:10.1016/j.quascirev.2011.07.002.
- Guthrie, R. Dale: Mammals of the mammoth steppe as paleoenvironmental indicators. In: Hopkins, D.M., Schweger, C.E., Young, S.B. (ed.): Paleoecology of Beringia. Academic Press, New York 1982, pp. 307–329.
- "Major Divisions". Subcommission on Quaternary Stratigraphy. International Commission on Stratigraphy. 4 January 2016. Retrieved 25 January 2017.
- Intergovernmental Panel on Climate Change (UN). "IPCC Fourth Assessment Report: Climate Change 2007 – Palaeoclimatic Perspective". The Nobel Foundation. Archived from the original on 2015-10-30. Retrieved 2016-03-28.
- "Woolly mammoth diet mystery solved by DNA analysis".
- Clark, P. U.; Dyke, A. S.; Shakun, J. D.; Carlson, A. E.; Clark, J.; Wohlfarth, B.; Mitrovica, J. X.; Hostetler, S. W.; McCabe, A. M. (2009). "The Last Glacial Maximum". Science. 325 (5941): 710–4. Bibcode:2009Sci...325..710C. doi:10.1126/science.1172873. PMID 19661421.
- Elias, Scott A.; Short, Susan K.; Nelson, C. Hans; Birks, Hilary H. (1996). "Life and times of the Bering land bridge". Nature. 382 (6586): 60–63. Bibcode:1996Natur.382...60E. doi:10.1038/382060a0.
- Hopkins, D.M., Matthews, J.V., Schweger, C.E., Young, S.B., (Eds.), 1982. Paleoecology of Beringia. Academic Press, New York.
- Vrba, E.S., Denton, G.H., Partridge, T.C., Buckle, L.H. (Eds.), 1995. Paleoclimate and Evolution With Emphasis on Human Origins. Yale University Press, New Haven.
- Nogués-Bravo, David; Rodríguez, Jesús; Hortal, Joaquín; Batra, Persaram; Araújo, Miguel B (2008). "Climate Change, Humans, and the Extinction of the Woolly Mammoth". PLoS Biology. 6 (4): e79. doi:10.1371/journal.pbio.0060079. PMC 2276529. PMID 18384234.
- Dale Guthrie, R. (2006). "New carbon dates link climatic change with human colonization and Pleistocene extinctions". Nature. 441 (7090): 207–9. Bibcode:2006Natur.441..207D. doi:10.1038/nature04604. PMID 16688174.
- Van Geel, Bas; Aptroot, André; Baittinger, Claudia; Birks, Hilary H.; Bull, Ian D.; Cross, Hugh B.; Evershed, Richard P.; Gravendeel, Barbara; Kompanje, Erwin J.O.; Kuperus, Peter; Mol, Dick; Nierop, Klaas G.J.; Pals, Jan Peter; Tikhonov, Alexei N.; Van Reenen, Guido; Van Tienderen, Peter H. (2008). "The ecological implications of a Yakutian mammoth's last meal" (PDF). Quaternary Research. 69 (3): 361–376. Bibcode:2008QuRes..69..361V. doi:10.1016/j.yqres.2008.02.004.
- Keith Kirby and Charles Watkins (2015). "The Forest Landscape before Farming". Europe's Changing Woods and Forests: From Wildwood to Managed Landscapes. CAB International. p. 34. ISBN 9781780643373.CS1 maint: uses authors parameter (link)
- "Woolly mammoths died for want of a few herbs".
- Andreev, Andrei A.; Siegert, Christine; Klimanov, Vladimir A.; Derevyagin, Aleksandr Yu.; Shilova, Galina N.; Melles, Martin (2002). "Late Pleistocene and Holocene Vegetation and Climate on the Taymyr Lowland, Northern Siberia" (PDF). Quaternary Research. 57 (1): 138–150. Bibcode:2002QuRes..57..138A. doi:10.1006/qres.2001.2302.
- Bocherens, Hervé (June 2015). "Isotopic tracking of large carnivore palaeoecology in the mammoth steppe". Quaternary Science Reviews. 117: 42–71. doi:10.1016/j.quascirev.2015.03.018. ISSN 0277-3791.
- Garrut, N.V., Boeskorov, G.G., 2001. In: Rozanov, Yu.A. (Ed.), Mammoth and Its Environment: 200 Years of Investigations. GEOS, Moscow.
- Kuzmin, Y.A., Orlova, L.A., Zolnikov, I.D., Igolnikov, A.E., 2001. In: Rozanov, A.Yu. (Ed.), Mammoth and Its Environment: 200 Years of Investigations. GEOS, Moscow, pp. 124e138.
- Syroechkovskii, V.E., 1986. Severnii Olen’ Agropromizdat. Moscow (in Russian).
- Veltre, D. W.; Yesner, D. R.; Crossen, K. J.; Graham, R. W.; Coltrain, J. B. (2008). "Patterns of faunal extinction and paleoclimatic change from mid-Holocene mammoth and polar bear remains, Pribilof Islands, Alaska". Quaternary Research. 70 (1): 40–50. Bibcode:2008QuRes..70...40V. doi:10.1016/j.yqres.2008.03.006.
- Enk, J. M.; Yesner, D. R.; Crossen, K. J.; Veltre, D. W.; O'Rourke, D. H. (2009). "Phylogeographic analysis of the mid-Holocene Mammoth from Qagnaxˆ Cave, St. Paul Island, Alaska" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 273 (1–2): 184–190. Bibcode:2009PPP...273D...5.. doi:10.1016/j.palaeo.2008.12.019.
- Tikhonov, Alexei; Larry Agenbroad; Sergey Vartanyan (2003). "Comparative analysis of the mammoth populations on Wrangel Island and the Channel Islands". Deinsea. 9: 415–420. ISSN 0923-9308. Archived from the original on 2012-06-11.
- Arslanov, K., Cook, G.T. , Gulliksen, S., Harkness, D.D., Kankainen, T., Scott, E.M., Vartanyan, S., and Zaitseva, G.I. (1998). "Consensus Dating of Remains from Wrangel Island". Radiocarbon. 40 (1): 289–294. doi:10.1017/S0033822200018166. Retrieved 2012-03-07.CS1 maint: multiple names: authors list (link)
- Vartanyan, S. L.; Arslanov, Kh. A.; Tertychnaya, T. V.; Chernov, S. B. (1995). "Radiocarbon Dating Evidence for Mammoths on Wrangel Island, Arctic Ocean, until 2000 BC". Radiocarbon. 37 (1): 1–6. doi:10.1017/S0033822200014703. Retrieved 2008-01-10.
- Boeskorov, Gennady G.; Potapova, Olga R.; Protopopov, Albert V.; Plotnikov, Valery V.; Agenbroad, Larry D.; Kirikov, Konstantin S.; Pavlov, Innokenty S.; Shchelchkova, Marina V.; Belolyubskii, Innocenty N.; Tomshin, Mikhail D.; Kowalczyk, Rafal; Davydov, Sergey P.; Kolesov, Stanislav D.; Tikhonov, Alexey N.; Van Der Plicht, Johannes (2016). "The Yukagir Bison: The exterior morphology of a complete frozen mummy of the extinct steppe bison, Bison priscus from the early Holocene of northern Yakutia, Russia". Quaternary International. 406: 94–110. Bibcode:2016QuInt.406...94B. doi:10.1016/j.quaint.2015.11.084.
- Rabanus-Wallace, M. Timothy; Wooller, Matthew J.; Zazula, Grant D.; Shute, Elen; Jahren, A. Hope; Kosintsev, Pavel; Burns, James A.; Breen, James; Llamas, Bastien; Cooper, Alan (2017). "Megafaunal isotopes reveal role of increased moisture on rangeland during late Pleistocene extinctions". Nature Ecology & Evolution. 1 (5): 0125. doi:10.1038/s41559-017-0125. PMID 28812683.
- Pitulko, V. V.; Tikhonov, A. N.; Pavlova, E. Y.; Nikolskiy, P. A.; Kuper, K. E.; Polozov, R. N. (2016). "Early human presence in the Arctic: Evidence from 45,000-year-old mammoth remains". Science. 351 (6270): 260–3. Bibcode:2016Sci...351..260P. doi:10.1126/science.aad0554. PMID 26816376.
- Pitulko, V.V.; Pavlova, E.Y.; Basilyan, A.E. (2016). "Mass accumulations of mammoth (mammoth 'graveyards') with indications of past human activity in the northern Yana-Indighirka lowland, Arctic Siberia". Quaternary International. 406: 202–217. Bibcode:2016QuInt.406..202P. doi:10.1016/j.quaint.2015.12.039.
- Frenzel B, Pécsi M, Velichko AA (1992) Atlas of paleoclimates and paleoenvironments of the Northern Hemisphere. JenaNew York: Geographical Research Institute, Hungarian Academy of Sciences Budapest and Gustav Fischer Verlag Stuttgart. 153 p
- Agadjanian AK, Serdyuk NV (2005) The history of mammalian communities and paleogeography of the Altai Mountains in the Paleolithic. Paleontological Journal 39: 645–821.
- Tarasov, PE; Guiot, J; Cheddadi, R; Andreev, AA; Bezusko, LG; et al. (1999). "Climate in northern Eurasia 6000 years ago reconstructed from pollen data". Earth Planet Sci Lett. 171: 635–645. doi:10.1016/s0012-821x(99)00171-5.
- Tarasov, PE; Volkova, VS; Webb, III (2000). "T, Guiot J, Andreev AA, et al. (2000) Last glacial maximum biomes reconstructed from pollen and plant macrofossil data from northern Eurasia". J Biogeogr. 27: 609–620. doi:10.1046/j.1365-2699.2000.00429.x.
- Jankovská, V; Pokorný, P (2008). "Forest vegetation of the last full-glacial period in the Western Carpathians (Slovakia and Czech Republic)". Preslia. 80: 307–324.
- Kuneš, P; Pelánková, B; Chytrý, M; Jankovská, V; Pokorný, P; et al. (2008). "Interpretation of the last-glacial vegetation of eastern-central Europe using modern analogues from southern Siberia". J Biogeogr. 35: 2223–2236. doi:10.1111/j.1365-2699.2008.01974.x.
- Pelánková, B; Chytrý, M (2009). "Surface pollen-vegetation relationships in the forest-steppe, taiga and tundra landscapes of the Russian Altai Mountains". Rev Palaeobot Palyno. 157: 253–265. doi:10.1016/j.revpalbo.2009.05.005.