Carbamoyl phosphate synthetase I
Carbamoyl phosphate synthetase I is a ligase enzyme located in the mitochondria involved in the production of urea. Carbamoyl phosphate synthetase I (CPS1 or CPSI) transfers an ammonia molecule from glutamine or glutamate to a molecule of bicarbonate that has been phosphorylated by a molecule of ATP. The resulting carbamate is then phosphorylated with another molecule of ATP. The resulting molecule of carbamoyl phosphate leaves the enzyme.
| carbamoyl-phosphate synthetase 1, mitochondrial | |
|---|---|
| Identifiers | |
| Symbol | CPS1 |
| NCBI gene | 1373 |
| HGNC | 2323 |
| OMIM | 608307 |
| RefSeq | NM_001875 |
| UniProt | P31327 |
| Other data | |
| EC number | 6.3.4.16 |
| Locus | Chr. 2 p |
Structure
In E. coli CPSI is a heterodimer with a small subunit and a larger subunit with about 382 and 1073 amino acid residues in size, although in mammals (and other vertebrates) the CPSI protein is encoded by a single gene.[1] The small subunit contains one active site for the binding and deamination of glutamine to make ammonia and glutamate. The large subunit contains two active sites, one for the production of carboxyphosphate, and the other for the production of carbamoyl phosphate.[2][3] Within the large subunit there are two domains (B and C) each with an active site of the ATP-grasp family.[1] Connecting the two subunits is a tunnel of sorts, which directs the ammonia from the small subunit to the large subunit.[4]
Mechanism
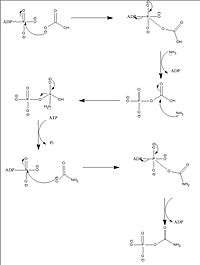
The overall reaction that occurs in CPSI is:
2ATP + HCO3− + NH4+ → 2ADP + Carbamoyl phosphate + Pi[4]
This reaction can be thought of occurring in four distinct steps.[5]
- Bicarbonate is phosphorylated
- Ammonia is cleaved from glutamine (glutaminase) or glutamate (glutamate dehydrogenase)
- The ammonia attacks the carboxyphosphate, resulting in carbamate
- Carbamate is phosphorylated to give Carbamoyl phosphate
Of these four steps, only step two—the deamination of glutamine to get ammonia—is known to have actively participating amino acid residues, Cys269 and His353. The other three steps mostly utilize amino acid residue to form hydrogen bonds with substrates. A video of a simplified version of this mechanism is available here.
Recent mechanism studies
It has been found that both ATP-binding sites in the large subunit of CPSI are structurally equivalent. A recent study has investigated the interlinking between these two domains (domain B and domain C) and has found evidence that they are coupled. This ATP-binding domain coupling works in a way such that a molecule of ATP binding at one site (domain C) conformationally allows synthesis at the other domain (domain B). If this is the case, carbamoyl phosphate is, in fact, not formed in step 5 (of the included mechanism below) by ejecting ADP but rather in step 4 by protonating the alcohol group and then kicking it off as water.[6]
Regulation
CPSI is regulated by N-acetylglutamate which acts as an obligate allosteric activator of CPS1. NAG, by binding to domain L4, triggers changes in the A-loop and in Arg1453 that result in changing interactions with the T′-loop of domain L3, which reorganizes completely from a β-hairpin in the apo form to a widened loop in the ligand-bound form. In this last form, the T′-loop interacts also with the tunnel-loop and the T-loop of the L1 domain, thus transferring the activating information to the bicarbonate-phosphorylating domain. This interaction with NAG and a second interaction, with a nucleotide, stabilise the active form of CPSI.[n 1] The necessity for this ligand also connects the high concentration of nitrogen, reflected in excess of glutamate and arginine to produce NAG, to an increase in CPSI activity to clear this excess.
Metabolism
CPSI plays a vital role in protein and nitrogen metabolism. Once ammonia has been brought into the mitochondria via glutamine or glutamate, it is CPSI's job to add the ammonia to bicarbonate along with a phosphate group to form carbamoyl phosphate. Carbamoyl phosphate is then put into the urea cycle to eventually create urea. Urea can then be transferred back to the blood stream and to the kidneys for filtration and on to the bladder for excretion.[8]
Related health problems
The main problem related to CPSI is genetics-based. Sometimes the body does not produce enough CPSI due to a mutation in the genetic code, resulting in poor metabolism of proteins and nitrogen, as well as high levels of ammonia in the body. This is dangerous because ammonia is highly toxic to the body, especially the nervous system, and can result in retardation and seizures.
Notes
- de Cima S, Polo LM, Díez-Fernández C, Martínez AI, Cervera J, Fita I, Rubio V (November 2015). "Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis". Scientific Reports. 5 (1): 16950. doi:10.1038/srep16950. PMC 4655335. PMID 26592762.
References
- Thoden JB, Huang X, Raushel FM, Holden HM (October 2002). "Carbamoyl-phosphate synthetase. Creation of an escape route for ammonia". The Journal of Biological Chemistry. 277 (42): 39722–7. doi:10.1074/jbc.M206915200. PMID 12130656.
- Powers SG, Griffith OW, Meister A (May 1977). "Inhibition of carbamyl phosphate synthetase by P1, P5-di(adenosine 5')-pentaphosphate: evidence for two ATP binding sites". The Journal of Biological Chemistry. 252 (10): 3558–60. PMID 193838.
- Thoden JB, Holden HM, Wesenberg G, Raushel FM, Rayment I (May 1997). "Structure of carbamoyl phosphate synthetase: a journey of 96 A from substrate to product". Biochemistry. 36 (21): 6305–16. doi:10.1021/bi970503q. PMID 9174345.
- Kim J, Raushel FM (May 2004). "Perforation of the tunnel wall in carbamoyl phosphate synthetase derails the passage of ammonia between sequential active sites". Biochemistry. 43 (18): 5334–40. doi:10.1021/bi049945+. PMID 15122899.
- Meister A (1989). "Mechanism and Regulation of the Glutamine-Dependent Carbamyl Phosphate Synthetase of Escherichia Coli". Mechanism and regulation of the glutamine-dependent carbamyl phosphate synthetase of Escherichia coli. Advances in Enzymology and Related Areas of Molecular Biology. 62. pp. 315–74. doi:10.1002/9780470123089.ch7. ISBN 9780470123089. PMID 2658488.
- Kothe M, Eroglu B, Mazza H, Samudera H, Powers-Lee S (November 1997). "Novel mechanism for carbamoyl-phosphate synthetase: a nucleotide switch for functionally equivalent domains". Proceedings of the National Academy of Sciences of the United States of America. 94 (23): 12348–53. doi:10.1073/pnas.94.23.12348. PMC 24940. PMID 9356452.
- Thoden JB, Miran SG, Phillips JC, Howard AJ, Raushel FM, Holden HM (June 1998). "Carbamoyl phosphate synthetase: caught in the act of glutamine hydrolysis". Biochemistry. 37 (25): 8825–31. doi:10.1021/bi9807761. PMID 9636022.
- Nelson D, Cox M. Principles of Biochemistry (fourth ed.). pp. 666–669.
External links
- GeneReviews/NCBI/NIH/UW entry on Urea Cycle Disorders Overview
- Carbamoyl-Phosphate+Synthase+(Ammonia) at the US National Library of Medicine Medical Subject Headings (MeSH)