Meganisoptera
Meganisoptera is an extinct order of very large to gigantic insects, occasionally called griffinflies. The order was formerly named Protodonata, the "proto-Odonata", for their similar appearance and supposed relation to modern Odonata (damselflies and dragonflies). They range in Palaeozoic (Late Carboniferous to Late Permian) times. Though most were only slightly larger than modern dragonflies, the order includes the largest known insect species, such as the late Carboniferous Meganeura monyi, Megatypus, and the even larger early Permian Meganeuropsis permiana, with wingspans of up to 71 centimetres (28 in).[1]

| Meganisoptera | |
|---|---|
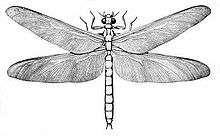 | |
| Illustration of a Meganeura species | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Superorder: | Odonatoptera |
| Order: | †Meganisoptera Martynov, 1932 |
| Families | |
| |
| Synonyms | |
| |
The forewings and hindwings are similar in venation (a primitive feature) except for the larger anal (rearwards) area in the hindwing. The forewing is usually more slender and slightly longer than the hindwing. Unlike the true dragonflies, the Odonata, they had no pterostigma, and a somewhat simpler pattern of veins in the wings.
Most specimens are known from wing fragments only; with only a few as complete wings, and even fewer (of the family Meganeuridae) with body impressions. These show a globose head with large dentate mandibles, strong spiny legs, a large thorax, and long and slender dragonfly-like abdomen. Like true dragonflies, they were presumably predators.
A few nymphs are also known, and show mouthparts similar to those of modern dragonfly nymphs, suggesting that they were also active aquatic predators.[3]
Although sometimes included under the dragonflies, the Protodonata lack certain distinctive wing features that characterise the Odonata. Grimaldi & Engel 2005 point out that the colloquial term "giant dragonfly" is therefore misleading, and suggest "griffinfly" instead.
Size

Controversy has prevailed as to how insects of the Carboniferous period were able to grow so large. The way oxygen is diffused through the insect's body via its tracheal breathing system puts an upper limit on body size, which prehistoric insects seem to have well exceeded. It was originally proposed in Harlé (1911) that Meganeura was only able to fly because the atmosphere at that time contained more oxygen than the present 20%. This theory was dismissed by fellow scientists, but has found approval more recently through further study into the relationship between gigantism and oxygen availability.[4] If this theory is correct, these insects would have been susceptible to falling oxygen levels and certainly could not survive in our modern atmosphere. Other research indicates that insects really do breathe, with "rapid cycles of tracheal compression and expansion".[5] Recent analysis of the flight energetics of modern insects and birds suggests that both the oxygen levels and air density provide a bound on size.[6]
A general problem with all oxygen related explanations of giant griffinflies is the circumstance that very large Meganeuridae with a wingspan of 45 cm also occurred in the Upper Permian of Lodève in France, when the oxygen content of the atmosphere was already much lower than in the Carboniferous and Lower Permian.[7]
Bechly 2004 suggested that the lack of aerial vertebrate predators allowed pterygote insects to evolve to maximum sizes during the Carboniferous and Permian periods, maybe accelerated by an "evolutionary arms race" for increase in body size between plant-feeding Palaeodictyoptera and meganeurids as their predators.
Families and genera
These families belong to the order Meganisoptera:[8]
- Aulertupidae Zessin & Brauckmann 2010
- Kohlwaldiidae Guthörl 1962
- Meganeuridae Handlirsch 1906
- Namurotypidae Bechly 1996
- Paralogidae Handlirsch 1906
These genera belong to the order Meganisoptera, but have not been placed in families:[8]
- Alanympha Kukalova-Peck 2009
- Asapheneura Pruvost 1919
- Dragonympha Kukalova-Peck 2009
- Palaeotherates Handlirsch 1906
- Paralogopsis Handlirsch 1911
- Schlechtendaliola Handlirsch, 1919
- Typoides Zalessky 1948
Notes
- The model in this photograph incorrectly depicts pterostigma on the wings.
References
- Grimaldi & Engel 2005, p. 175.
- Tillyard 1917, p. 324: "No Dragonfly at present existing can compare with the immense Meganeura monyi of the Upper Carboniferous, whose expanse of wing was somewhere about twenty-seven inches."
- Hoell, Doyen & Purcell 1998, p. 321.
- Chapelle & Peck 1999, pp. 114–115: "Oxygen supply may also have led to insect gigantism in the Carboniferous period, because atmospheric oxygen was 30-35% (ref. 7). The demise of these insects when oxygen content fell indicates that large species may be susceptible to such change. Giant amphipods may therefore be among the first species to disappear if global temperatures are increased or global oxygen levels decline. Being close to the critical MPS limit may be seen as a specialization that makes giant species more prone to extinction over geological time."
- Westneat et al. 2003: "Insects are known to exchange respiratory gases in their system of tracheal tubes by using either diffusion or changes in internal pressure that are produced through body motion or hemolymph circulation. However, the inability to see inside living insects has limited our understanding of their respiration mechanisms. We used a synchrotron beam to obtain x-ray videos of living, breathing insects. Beetles, crickets, and ants exhibited rapid cycles of tracheal compression and expansion in the head and thorax. Body movements and hemolymph circulation cannot account for these cycles; therefore, our observations demonstrate a previously unknown mechanism of respiration in insects analogous to the inflation and deflation of vertebrate lungs."
- Dudley 1998: "Uniformitarian approaches to the evolution of terrestrial locomotor physiology and animal flight performance have generally presupposed the constancy of atmospheric composition. Recent geophysical data as well as theoretical models suggest that, to the contrary, both oxygen and carbon dioxide concentrations have changed dramatically during defining periods of metazoan evolution. Hyperoxia in the late Paleozoic atmosphere may have physiologically enhanced the initial evolution of tetrapod locomotor energetics; a concurrently hyperdense atmosphere would have augmented aerodynamic force production in early flying insects. Multiple historical origins of vertebrate flight also correlate temporally with geological periods of increased oxygen concentration and atmospheric density. Arthropod as well as amphibian gigantism appear to have been facilitated by a hyperoxic Carboniferous atmosphere and were subsequently eliminated by a late Permian transition to hypoxia. For extant organisms, the transient, chronic and ontogenetic effects of exposure to hyperoxic gas mixtures are poorly understood relative to contemporary understanding of the physiology of oxygen deprivation. Experimentally, the biomechanical and physiological effects of hyperoxia on animal flight performance can be decoupled through the use of gas mixtures that vary in density and oxygen concentration. Such manipulations permit both paleophysiological simulation of ancestral locomotor performance and an analysis of maximal flight capacity in extant forms."
- Nel et al. 2008.
- "The Paleobiology Database, order Meganisoptera". Retrieved 2019-10-17.
Bibliography
- Bechly, G (2004). "Evolution and systematics" (PDF). In Hutchins, M.; Evans, A.V.; Garrison, R.W. & Schlager, N. (eds.). Grzimek's Animal Life Encyclopedia. Insects (2nd ed.). Farmington Hills, MI: Gale. pp. 7–16.CS1 maint: ref=harv (link)
- Carpenter, F. M. (1992). "Superclass Hexapoda". Treatise on Invertebrate Paleontology. Volume 3 of Part R, Arthropoda 4. Boulder, CO: Geological Society of America.CS1 maint: ref=harv (link)
- Dudley, Robert (April 1998). "Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotion performance". The Journal of Experimental Biology. 201 (Pt8): 1043–1050. PMID 9510518.CS1 maint: ref=harv (link)
- Chapelle, Gauthier & Peck, Lloyd S. (May 1999). "Polar gigantism dictated by oxygen availability". Nature. 399 (6732): 114–115. doi:10.1038/20099.CS1 maint: ref=harv (link)
- Grimaldi, David & Engel, Michael S. (2005-05-16). Evolution of the Insects. Cambridge University Press. ISBN 978-0-521-82149-0.CS1 maint: ref=harv (link)
- Harlé, Edouard (1911). "Le Vol de grands reptiles et insectes disparus semble indiquer une pression atmosphérique élevée". Extr. Du Bulletin de la Sté Géologique de France (in French). 4 (9): 118–121.CS1 maint: ref=harv (link)
- Hoell, H.V.; Doyen, J.T. & Purcell, A.H. (1998). Introduction to Insect Biology and Diversity (2nd ed.). Oxford University Press. ISBN 978-0-19-510033-4.CS1 maint: ref=harv (link)
- Nel, André; Fleck, Günther; Garrouste, Romain; Gand, Georges; Lapeyrie, Jean; Bybee, Seth M & Prokop, Jakub (2009). "Revision of Permo-Carboniferous griffenflies (Insecta: Odonatoptera: Meganisoptera) based upon new species and redescription of selected poorly known taxa from Eurasia". Palaeontographica Abteilung A. 289 (4–6): 89–121. doi:10.1127/pala/289/2009/89.CS1 maint: ref=harv (link)
- Nel, André; Fleck, Günther; Garrouste, Romain & Gand, Georges (2008). "The Odonatoptera of the Late Permian Lodève Basin (Insecta)". Journal of Iberian Geology. 34 (1): 115–122.CS1 maint: ref=harv (link)
- Tasch, Paul (1980) [1973]. Paleobiology of the Invertebrates. John Wiley and Sons. p. 617.CS1 maint: ref=harv (link)
- Tillyard, R.J. (1917). The Biology of Dragonflies. Cambridge University Press. p. 324. GGKEY:0Z7A1R071DD.CS1 maint: ref=harv (link)
- Westneat, MW; Betz, O; Blob, RW; Fezzaa, K; Cooper, WJ & Lee, WK (January 2003). "Tracheal respiration in insects visualized with synchrotron x-ray imaging". Science. 299 (5606): 558–560. doi:10.1126/science.1078008. PMID 12543973.CS1 maint: ref=harv (link)
External links
| Wikispecies has information related to Protodonata |
Insect orders | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
