Desmostylia
The Desmostylia (from Greek δεσμά desma, "bundle", and στῦλος stylos, "pillar")[1] are an extinct order of aquatic mammals that existed from the early Oligocene (Rupelian) to the late Miocene (Tortonian) (30.8 to 7.25 million years ago).
| Desmostylia | |
|---|---|
 | |
| Desmostylus, Royal Ontario Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Superorder: | Afrotheria |
| Order: | †Desmostylia Reinhart 1959 |
| Families and genera | |
| |
Desmostylians are the only known extinct order of marine mammals[2] (Thalassocnus, a genus of marine sloths, became extinct more recently).
The Desmostylia, together with Sirenia and Proboscidea (and possibly Embrithopoda), have traditionally been assigned to the afrotherian clade Tethytheria, a group named after the paleoocean Tethys around which they originally evolved. The relationship between the Desmostylia and the other orders within the Tethytheria has been disputed; if the common ancestor of all tethytheres was semiaquatic, the Proboscidea became secondarily terrestrial; alternatively, the Desmostylia and Sirenia could have evolved independently into aquatic mammals.[3] The assignment of Desmostylia to Afrotheria has always been problematic from a biogeographic standpoint, given that Africa was the locus of the early evolution of the Afrotheria while the Desmostylia have only been found along the Pacific Rim. That assignment has been seriously undermined by a 2014 cladistic analysis that places anthracobunids and desmostylians, two major groups of putative non-African afrotheres, close to each other within the laurasiatherian order Perissodactyla.[4] However, a posterior study shows that, while anthracobunids are definite perissodactyls, desmostylians share the same number of characteristics necessary for either Paenungulata or Perissodactyla, making their former assessment as afrotheres a possibility.[5]
Description

Desmostylians were large, fully aquatic quadrupeds with massive limbs and short tails.[2] The smallest is Ashoroa laticosta, a relatively large animal at a body length of 168 cm, while the largest species reached sizes comparable to Steller's sea cow.[6]
A desmostylian skull has an elongated and broadened rostrum, with the nasal opening located slightly dorsally. The zygomatic arches are prominent (behind the eyes), the paroccipital processes elongated (downward-pointing processes behind the jaw-joints), and the epitympanic sinuses open into the temporal fossae (cavities above the ear holes).[2]
The mandible and maxilla typically have forward-pointing incisors and canine tusks, followed by a long postcanine diastema, partly because of the reduced number of premolars. The cusps of the premolars and molars are composed of densely packed cylinders of thick enamel, giving the order its name ("bundle of columns"). The primitive dental formula is 3.1.4.3, with a trilobate fourth deciduous premolar. The cheek teeth are brachydont and bunodont in primitive genera, but hypsodont in later genera such as Desmostylus, which has many supernumerary cusps.[2]
In the postcrania, the clavicle is absent and the sternum consists of a series of heavy, paired, plate-like sternebrae. In adults, the joints between the radius and ulna prevent any movements. The metacarpals are longer than metatarsals, and each foot has four digits (digit I is vestigial).[2]
Behaviour
Their dental and skeletal forms suggest that desmostylians were aquatic herbivores dependent on littoral habitats. Their name refers to their highly distinctive molars, in which each cusp was modified into hollow columns, so a typical molar would have resembled a cluster of pipes, or in the case of worn molars, volcanoes. (This may reflect the close relationship between the Paenungulata, to which this group has been assigned, and the Tubulidentata.)
Desmostylus does not chew or eat like any other known animal. It clenches its teeth, roots up plants with the help of tusks and powerful neck, and then sucks them in using strong throat muscles and the shape of the roof of the mouth.[7]
Desmostylians are believed to be aquatic because of a combination of characteristics. Their legs seemed to be adapted for terrestrial locomotion, while a number of other parameters confirms their aquatic nature:[1]
- Fossils have been found in marine strata.
- The nares are retracted and the orbits are raised like in other aquatic mammals.
- Levels of stable isotopes in their tooth enamel suggest an aquatic diet and environment (carbon and oxygen) and fresh or brackish water (strontium).
- Their spongy bone structure is similar to that of cetaceans.
Based on a comparison of trunk and limb proportions, Gingerich 2005 concluded[8] that desmostylians were more terrestrial than aquatic and clearly fore limb-dominated swimmers, hence they were more similar to "sea bears" than "sea sloths" (as proposed by other researchers.) However, a more recent and detailed analysis of desmostylian bone structure has revealed them to be fully aquatic, like sirenians and cetaceans,[9] with their limbs being incapable of supporting their own weight on land. More recent studies vindicate this assessment, as desmostylians had a thoracic morphology more similar to sirenians and cetaceans than to that of semiaquatic mammals.[10] Its less dense bone structure suggests that Desmostylus had a lifestyle of active swimming and possibly feeding at the surface, while other desmostylians were primarily slow swimmers and/or bottom walkers and sea grass feeders.[9]
Habitat
A 2017 study on Desmostylus and Paleoparadoxia shows that the former preferred areas shallower than 30 m, while the latter occurred in deep, offshore waters.[11]
Distribution
Desmostylian fossils are known from the northern Pacific Rim,[12] from southern Japan through Russia, the Aleutian Islands, and the Pacific coast of North America to the southern tip of Baja California. They range from the Early Oligocene to the late Miocene.
Extinction
Desmostylians, being fully marine herbivores, are thought to have been outcompeted ecologically by dugongid sirenians. In particular, later species like Neoparadoxia are more specialised than previous forms, suggesting increased divergence to compete with sirenians, and sirenian diversity appears to increase with desmostylian decline.[13] Both desmostylians and North Pacific dugongids were apparently kelp specialists, as opposed to marine herbivorous mammals from other regions, with diets primarily composed of seagrass.[6]
Classification
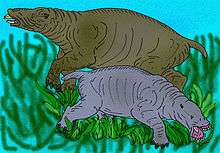



The type species Desmostylus hesperus was originally classified from a few teeth and vertebrae as a sirenian by Marsh 1888, but doubts arose a decade later when more complete fossils were discovered in Japan. Osborn 1905 also proposed that they belonged to the Sirenia.[14] One of the most comprehensive collections of desmostylian teeth was amassed by paleontologist John C. Merriam, who concluded on the basis of the molar structure and repeated occurrence in marine beds that the animals had been aquatic, and were probably sirenian.
In 1926, the Austrian palaeontologist Othenio Abel suggested origins with monotremes, like the duck-billed platypus, and in 1933, he even created the order "Desmostyloidea", which he placed within the Multituberculata. Abel died shortly after World War II, and his classification won few supporters and has been ignored since.[15]
Because desmostylians were originally known only from skull fragments, teeth, and bits of other bones, general agreement was that they had had flippers and a fin-like tail. The discovery of a complete skeleton from Sakhalin Island in 1941, however, showed that they possessed four legs, with bones as stout as a hippopotamus', and justified the creation of a new order for the desmostylians, described by Reinhart 1959.
A major find was announced in October 2015 after scientists examined an extensive group of giant, tusked, quadruped, marine mammal fossils. This northernmost to date species discovery had been unearthed during excavation for the construction of a school in Unalaska, in the Aleutian Islands.[16] A rendition of a group was drawn by Alaskan artist Ray Troll.[17]
Despite their similarities to manatees and elephants, desmostylians were entirely unlike any living creatures. Douglas Emlong's 1971 discovery of the new genus Behemotops from Oregon showed that early desmostylians had more proboscidean-like teeth and jaws than later ones. Despite this discovery, their relationships to manatees and proboscids remain unresolved. The analysis of Cooper et al. (2014) indicates the similarities with manatees and elephants may be a result of convergence and that they may instead be basal perissodactyls.[4]
Barnes 2013 proposed a new classification of Paleoparadoxiidae:[18]
- Order Desmostylia Reinhart, 1953
- Family Paleoparadoxiidae Reinhart, 1959
- Subfamily Behemotopsinae (Inuzuka, 1987)
- Behemotops Domning, Ray, and McKenna, 1986
- Behemotops proteus Domning, Ray, and McKenna, 1986 (including Behemotops emlongi Domning, Ray, and McKenna, 1986)
- Behemotops katsuiei Inuzuka, 2000b
- Behemotops Domning, Ray, and McKenna, 1986
- Subfamily Paleoparadoxiinae (Reinhart, 1959)
- Archaeoparadoxia
- Archaeoparadoxia weltoni (Clark, 1991)
- Paleoparadoxia Reinhart, 1959
- Paleoparadoxia tabatai (Tokunaga, 1939), (= Paleoparadoxia media Inuzuka, 2005)
- Neoparadoxia Barnes 2013
- Neoparadoxia repenningi (Domning and Barnes, 2007)
- Neoparadoxia cecilialina Barnes 2013
- Archaeoparadoxia
- Subfamily Behemotopsinae (Inuzuka, 1987)
- Family Paleoparadoxiidae Reinhart, 1959
References
- Gingerich 2005, Introduction
- Gheerbrant, Domning & Tassy 2005, pp. 95–6
- Uhen 2007, p. 515
- Cooper et al. 2014
- Gheerbrant, Emmanuel; Filippo, Andrea; Schmitt, Arnaud (2016). "Convergence of Afrotherian and Laurasiatherian Ungulate-Like Mammals: First Morphological Evidence from the Paleocene of Morocco". PLoS ONE. 11 (7): e0157556. doi:10.1371/journal.pone.0157556. PMC 4934866. PMID 27384169.
- Nicholas D. Pyenson, Geerat J. Vermeij, The rise of ocean giants: maximum body size in Cenozoic marine mammals as an indicator for productivity in the Pacific and Atlantic Oceans, Published 5 July 2016.DOI: 10.1098/rsbl.2016.0186
- "New fossils intensify mystery of short-lived, toothy mammal found in ancient North Pacific, Oddball creature, Desmostylia". www.sciencedaily.com. Southern Methodist University. Oct 6, 2015. Retrieved October 11, 2015.
- Gingerich 2005, Discussion
- Hayashi et al. 2013
- Ando, Konami (2016). "Farewell to life on land - thoracic strength as a new indicator to determine paleoecology in secondary aquatic mammals". Journal of Anatomy. 229: 768–777. doi:10.1111/joa.12518. PMC 5108153. PMID 27396988.
- Matsui, Kumiko; Sashida, Katsuo; Agematsu, Sachiko; Kohno, Naoki (2017). "Habitat preferences of the enigmatic Miocene tethythere Desmostylus and Paleoparadoxia (Desmostylia; Mammalia) inferred from the depositional depth of fossil occurrences in the Northwestern Pacific realm". Palaeogeography, Palaeoclimatology, Palaeoecology. 471: 254–265. doi:10.1016/j.palaeo.2017.02.005.
- Gingerich 2005, Abstract
- Barnes, L.. (2013, September 11) .A new genus and species of late Miocene Paleoparadoxiid (Mammalia, Desmostylia) from California. Natural History Museum of Los Angeles County Contributions in Science, (521), 51–114
- H. F. Osborn 1905 in the Paleobiology Database. Retrieved March 2013.
- Domning, Ray & McKenna 1986, p. 34
- Giant extinct mammal identified from Unalaska fossils, Alaska Dispatch News, Mike Dunham, October 7, 2015. Retrieved 8 October 2015.
- Photos: New species of extinct marine mammal identified from Unalaska fossils, Alaska Dispatch News, Mike Dunham, October 7, 2015. Retrieved 8 October 2015.
- Barnes 2013, pp. 103, 108–109
Further reading
- Barnes, Lawrence G. (2013). "A New Genus of Late Miocene Paleoparadoxiid (Mammalia, Desmostylia) from California" (PDF). Contributions in Science. 521: 51–114. Retrieved 14 January 2014.CS1 maint: ref=harv (link)
- Cooper, L. N.; Seiffert, E. R.; Clementz, M.; Madar, S. I.; Bajpai, S.; Hussain, S. T.; Thewissen, J. G. M. (2014-10-08). "Anthracobunids from the Middle Eocene of India and Pakistan Are Stem Perissodactyls". PLoS ONE. 9 (10): e109232. doi:10.1371/journal.pone.0109232. PMC 4189980. PMID 25295875.CS1 maint: ref=harv (link)
- Domning, Daryl P.; Barnes, Lawrence G. (2007). "A new name for the 'Stanford skeleton' of Paleoparadoxia (Mammlia, Desmostylia)". Journal of Vertebrate Paleontology. 27 (3): 748–751. doi:10.1671/0272-4634(2007)27[748:annfts]2.0.co;2.CS1 maint: ref=harv (link)
- Domning, Daryl P.; Ray, Clayton Edward; McKenna, Malcolm C. (1986). "Two new Oligocene desmostylians and a discussion of Tethytherian systematics" (PDF). Smithsonian Contributions to Paleobiology. 59 (59): 1–56. doi:10.5479/si.00810266.59.1. OCLC 12974759. Retrieved 15 December 2013.CS1 maint: ref=harv (link)
- Gingerich, Philip D. (2005). "Aquatic Adaptation and Swimming Mode Inferred from Skeletal Proportions in the Miocene Desmostylian Desmostylus" (PDF). Journal of Mammalian Evolution. 12 (1/2): 183–194. doi:10.1007/s10914-005-5719-1. Retrieved March 2013. Check date values in:
|accessdate=(help)CS1 maint: ref=harv (link) - Hay, O. P. (1923). "Characteristics of sundry fossil vertebrates". Pan-American Geologist. 39: 101–20. OCLC 38855496.CS1 maint: ref=harv (link)
- Hayashi, S.; Houssaye, A.; Nakajima, Y.; Kentaro, C.; Ando, T.; Sawamura, H.; Inuzuka, N.; Kaneko, N.; Osaki, T. (2013). "Bone Inner Structure Suggests Increasing Aquatic Adaptations in Desmostylia (Mammalia, Afrotheria)". PLoS ONE. 8 (4): e59146. doi:10.1371/journal.pone.0059146. OCLC 837402105. PMC 3615000. PMID 23565143.CS1 maint: ref=harv (link)
- Inuzuka, N. (2000). "Primitive late Oligocene desmostylians from Japan and Phylogeny of the Desmostylia". Bulletin of the Ashoro Museum of Paleontology. 1: 91–123. Lay summary (December 2013).CS1 maint: ref=harv (link)
- Marsh, O. C. (1888). "Notice of a new fossil sirenian, from California" (PDF). American Journal of Science. 25 (8): 94–96. OCLC 79838746. Retrieved 15 December 2013.CS1 maint: ref=harv (link)
- Osborn, H. F. (1905). "Ten years progress in the mammalian Palaeontology of North America". Extrait des Comptes Rendus du 6e Congres International de Zoologie. Session de Berne 1904: 86–113. OCLC 18502464.CS1 maint: ref=harv (link)
- Pronina, I. G. (1957). "A new desmostylid, Kronokotherium brevimaxillare gen. nov., sp. nov., from the Miocene deposits of Kamchatka". C. R. Acad. Sci. URSS. 117: 310–12.CS1 maint: ref=harv (link)
- Reinhart, Roy Herbert (1959). "A review of the Sirenia and Desmostylia". University of California Publications in Geological Sciences. 36 (1): 1–146. OCLC 3474601.CS1 maint: ref=harv (link)
- Gheerbrant, Emmanuel; Domning, Daryl P.; Tassy, Pascal (2005). "Paenungulata". In Rose, Kenneth D.; Archibald, J. David (eds.). The Rise of Placental Mammals: Origins and Relationships of the Major Extant Clades. JHU Press. ISBN 9780801880223. OCLC 55801049. Retrieved June 2013. Check date values in:
|accessdate=(help)CS1 maint: ref=harv (link) - Uhen, Mark D. (2007). "Evolution of Marine Mammals: Back to the Sea After 300 Million Years". Anatomical Record. 290 (6): 514–22. doi:10.1002/ar.20545. OCLC 137264359. PMID 17516441.CS1 maint: ref=harv (link)
External links
| Wikispecies has information related to Desmostylia |
| Look up desmostylia in Wiktionary, the free dictionary. |
| Wikimedia Commons has media related to Desmostylia. |
- "Introduction to Desmostylia". University of California Museum of Paleontology. January 2009. Retrieved 16 June 2013.