Fluorinase
The fluorinase enzyme (EC 2.5.1.63, also known as adenosyl-fluoride synthase) catalyzes the reaction between fluoride ion and the co-factor S-adenosyl-L-methionine to generate L-methionine and 5'-fluoro-5'-deoxyadenosine, the first committed product of the fluorometabolite biosynthesis pathway.[1] The fluorinase was originally isolated from the soil bacterium Streptomyces cattleya, but homologues have since been identified in a number of other bacterial species, including Streptomyces sp. MA37, Nocardia brasiliensis and Actinoplanes sp. N902-109.[2] This is the only known enzyme capable of catalysing the formation of a carbon-fluorine bond, the strongest single bond in organic chemistry.[3]

| Fluorinase (adenosyl-fluoride synthase) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 | |||||||||
| Identifiers | |||||||||
| EC number | 2.5.1.63 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
A homologous chlorinase enzyme, which catalyses the same reaction with chloride rather than fluoride ion, has been isolated from Salinospora tropica, from the biosynthetic pathway of salinosporamide A.[4]
Reactivity
The fluorinase catalyses an SN2-type nucleophilic substitution at the C-5' position of SAM, while L-methionine acts as a neutral leaving group.[5][6] The fluorinase-catalysed reaction is estimated to be between 106[6] to 1015[7] times faster than the uncatalysed reaction, a significant rate enhancement. Despite this, the fluorinase is still regarded as a slow enzyme, with a turnover number (kcat) of 0.06 min−1.[8] The high kinetic barrier to reaction is attributed to the strong solvation of fluoride ion in water, resulting in a high activation energy associated with stripping solvating water molecules from aqueous fluoride ion, converting fluoride into a potent nucleophile within the active site.
The reaction catalysed by the fluorinase is reversible, and upon incubation of 5'-fluoro-5'-deoxyadenosine and L-methionine with the fluorinase, SAM and fluoride ion are produced.[9] Replacing L-methionine with L-selenomethionine results in a 6-fold rate enhancement of the reverse reaction,[9] due to the increased nucleophilicity of the selenium centre compared to the sulfur centre.
The fluorinase shows a degree of substrate tolerance for halide ion, and can also use chloride ion in place of fluoride ion. While the equilibrium for reaction between SAM and fluoride ion lies towards products FDA and L-methionine, the equilibrium position is reversed in the case for chloride ion. Incubation of SAM and chloride ion with the fluorinase does not result in generation of 5'-chloro-5'-deoxyadenosine (ClDA), unless an additional enzyme, an L-amino acid oxidase, is added. The amino acid oxidase removes the L-methionine from the reaction, converting it to the corresponding oxo-acid.
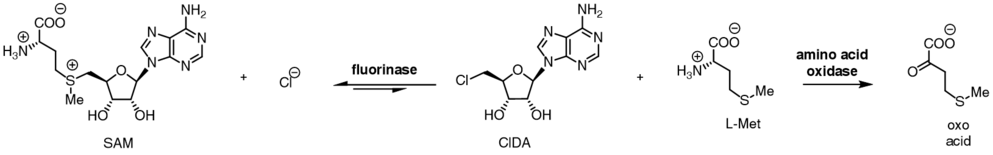
The halide preference, coupled to the position of the two reaction equilibria allows for a nett transhalogenation reaction to be catalysed by the enzyme.[9] Incubation of 5'-chloro nucleosides with the enzyme, along with catalytic L-selenomethionine or L-methionine results in the production of 5-fluoro nucleosides. When [18F]fluoride is used, this transhalogenation reaction can be used for the synthesis of radiotracers for positron emission tomography.[10][11][11]
Structural studies
As of late 2007, 9 structures have been solved for this class of enzymes, with PDB accession codes 1RQP, 1RQR, 2C2W, 2C4T, 2C4U, 2C5B, 2C5H, 2CBX, and 2CC2.
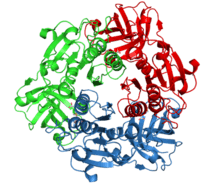
The names given to the enzyme come not from the structure, but from the function: 5-Fluoro-5-deoxyadenosine is the molecule synthesised. The structure is homologous to the duf-62 enzyme series. The enzyme is a dimer of trimers (2 molecules each with three subunits). The active sites are located between these subunits (subunit interfaces), each can bind to one SAM molecule at a time.[12]
Fluorometabolite biosynthesis
See also
References
- O'Hagan D, Schaffrath C, Cobb SL, Hamilton JT, Murphy CD (March 2002). "Biochemistry: biosynthesis of an organofluorine molecule". Nature. 416 (6878): 279. doi:10.1038/416279a. PMID 11907567.
- Deng H, Ma L, Bandaranayaka N, Qin Z, Mann G, Kyeremeh K, Yu Y, Shepherd T, Naismith JH, O'Hagan D (February 2014). "Identification of fluorinases from Streptomyces sp MA37, Norcardia brasiliensis, and Actinoplanes sp N902-109 by genome mining". ChemBioChem. 15 (3): 364–8. doi:10.1002/cbic.201300732. PMID 24449539.
- O'Hagan D (February 2008). "Understanding organofluorine chemistry. An introduction to the C-F bond". Chemical Society Reviews. 37 (2): 308–19. doi:10.1039/b711844a. PMID 18197347.
- Eustáquio AS, Pojer F, Noel JP, Moore BS (January 2008). "Discovery and characterization of a marine bacterial SAM-dependent chlorinase". Nature Chemical Biology. 4 (1): 69–74. doi:10.1038/nchembio.2007.56. PMC 2762381. PMID 18059261.
- Cadicamo CD, Courtieu J, Deng H, Meddour A, O'Hagan D (May 2004). "Enzymatic fluorination in Streptomyces cattleya takes place with an inversion of configuration consistent with an SN2 reaction mechanism". ChemBioChem. 5 (5): 685–90. doi:10.1002/cbic.200300839. PMID 15122641.
- Senn HM, O'Hagan D, Thiel W (October 2005). "Insight into enzymatic C-F bond formation from QM and QM/MM calculations". Journal of the American Chemical Society. 127 (39): 13643–55. doi:10.1021/ja053875s. PMID 16190730.
- Lohman DC, Edwards DR, Wolfenden R (October 2013). "Catalysis by desolvation: the catalytic prowess of SAM-dependent halide-alkylating enzymes". Journal of the American Chemical Society. 135 (39): 14473–5. doi:10.1021/ja406381b. PMID 24041082.
- Zhu X, Robinson DA, McEwan AR, O'Hagan D, Naismith JH (November 2007). "Mechanism of enzymatic fluorination in Streptomyces cattleya". Journal of the American Chemical Society. 129 (47): 14597–604. doi:10.1021/ja0731569. PMC 3326528. PMID 17985882.
- Deng H, Cobb SL, McEwan AR, McGlinchey RP, Naismith JH, O'Hagan D, Robinson DA, Spencer JB (January 2006). "The fluorinase from Streptomyces cattleya is also a chlorinase". Angewandte Chemie. 45 (5): 759–62. doi:10.1002/anie.200503582. PMC 3314195. PMID 16370017.
- Deng H, Cobb SL, Gee AD, Lockhart A, Martarello L, McGlinchey RP, O'Hagan D, Onega M (February 2006). "Fluorinase mediated C-(18)F bond formation, an enzymatic tool for PET labelling". Chemical Communications. 0 (6): 652–4. doi:10.1039/b516861a. PMID 16446840.
- Thompson S, Onega M, Ashworth S, Fleming IN, Passchier J, O'Hagan D (September 2015). "A two-step fluorinase enzyme mediated (18)F labelling of an RGD peptide for positron emission tomography". Chemical Communications. 51 (70): 13542–5. doi:10.1039/c5cc05013h. PMID 26221637.
- Dong C, Huang F, Deng H, Schaffrath C, Spencer JB, O'Hagan D, Naismith JH (February 2004). "Crystal structure and mechanism of a bacterial fluorinating enzyme". Nature. 427 (6974): 561–5. doi:10.1038/nature02280. PMID 14765200.