Dominance hierarchy
Dominance hierarchy is a type of social hierarchy that arises when members of a social group interact, to create a ranking system. In social living groups, members are likely to compete for access to limited resources and mating opportunities. Rather than fighting each time they meet, relative rank is established between members of the same sex. Based on repetitive interactions a social order is created that is subject to change each time a dominant animal is challenged by a subordinate one.[1]
Dominance
.JPG)
When individuals seek high rank
Given the benefits and costs of possessing a high rank within a hierarchal group, there are certain characteristics of individuals, groups, and environments that determine whether an individual will benefit from a high rank. Individual characteristics include whether or not high rank gives them access to valuable resources such as mates and food. Individuals will often weigh the cost of the resource against factors including their age, intelligence, experience, and physical fitness, which can determine the costs to gaining rank.
Hierarchy results as an accumulation of individual interaction, group dynamics, and sharing of resources, therefore group size and composition can affect the dominance decisions of high-ranking individuals and hierarchy type. For example, in a large group with many males, it may be very challenging for the highest-ranking male to dominate all the mating opportunities, so some mate sharing probably exists. These occasional mating opportunities available to subordinates reduce the likelihood of subordinates challenging the dominant male—mating is no longer an all-or-nothing game and those shared are enough to placate most subordinates. Another aspect that can determine dominance hierarchies is the environment. In populations of Kenyan vervets, high-ranking females have higher foraging success when the food resources are clumped, but when food is distributed throughout an area they lose their advantage because subordinate females can acquire food with less risk of encountering a dominant female.[2]
Benefits
Foraging success
Another benefit to high ranking individuals is increased foraging success and access to food resources. During times of water shortage the highest-ranking vervet females have greater access than subordinates females to water in tree holes. In chacma baboons, the high-ranking males have the first access to vertebrate prey that has been caught by the group, and in yellow baboons the dominant males feed for longer without being interrupted.
In many bird species the dominant individuals have higher rates of food intake including dark-eyed juncos and oystercatchers. The dominant individuals in these groups fill themselves up first and fill up more quickly, so they spend less time foraging, which reduces the risk of predation. Thus they have increased survival because of increased nutrition and decreased predation.[2]
Reproductive success
In primates, one of the most widely studied hierarchical groups, many studies have found a positive relationship between high rank and reproductive success. In baboons, higher-ranking males have the highest reproductive success due to increased female acquisition. Also, female baboons benefit from increased rank because high-ranking females produce more surviving offspring.[2] A 1991 meta-analysis of 32 studies determined there was a positive relationship between dominance rank and reproductive success amongst primates.[3] A 2016 study determined that higher status increased the reproductive success amongst men and the relationship between status and reproductive success did not depend upon the type of subsistence (forager, horticulture, pastoralism, agriculture), contrary to the "egalitarian hypothesis", which predicted status would have a weaker effect on reproductive success amongst foragers than amongst nonforagers.[4]
Bonnet macaques gain increased reproductive success with high rank. High-ranking males have more access to fertile females and consequently partake in most of the matings within the group; in one population, three males were responsible for over 75% of matings. In this population, males often vary in their rank, and as they gain rank, they gain more time spent exclusively with fertile females; the opposite is seen as males drop in rank.[5] In many primates, including bonnet macaques and rhesus monkeys, the offspring of high-ranking individuals have better fitness and thus an increased rate of survival. This is most likely a function of two factors. The first is that high-ranking males mate with high-ranking females. Assuming their high rank is correlated with higher fitness and fighting ability, this trait will be conferred to their offspring. The second factor is that higher-ranking parents probably provide better protection to their offspring and thus ensure higher survival rates.[2] Amongst rhesus macaques, higher ranking males sired more offspring, though the alpha male was never the one to sire the most offspring, with that instead being a high-ranking but not top male. The complex relationship between rank and reproduction in this species is likely explained by the fact that rhesus macaques queue, rather than fight, for dominance, meaning that the alpha male is not necessarily the strongest or most attractive male.[6][7]
In rodents, the highest-ranking male frequently sires the most offspring. The same pattern is found in most carnivores, such as the dwarf mongoose. The dwarf mongoose lives in a social system with one dominant pair. The dominant female produces all or almost all of the offspring in the living group, and the dominant male has first access to her during her oestrus period. In red deer, the males who experienced winter dominance, resulting from greater access to preferred foraging sites, had higher ability to get and maintain larger harems during the mating season.[2]
In many monogamous bird species, the dominant pairs tend to get the best territories, which in turn promote offspring survival and adult health. In dunnocks, a species of birds that experiences many mating systems, sometimes individuals will form a group that will have one dominant male who achieves all of the mating in the group.[2]
In the monogynous bee species Melipona subnitida it is noted that the queen wants to maintain reproductive success, and does so by preventing workers from caring for their cells, pushing or hitting them using her antennae. Workers additionally have been noted to display aggression towards males, claiming priority over the cells when males try to use them to place eggs. This species further demonstrates the conflict between hierarchy and reproductive success.[8][9]
Costs
Despite the benefits to being of a high rank in a hierarchal group, there are also costs which offset these benefits. The most common costs to high-ranking individuals are higher metabolic rates and higher levels of stress hormones.[2] In great tits and pied flycatchers, high-ranking individuals experience higher resting metabolic rates and therefore need to consume more food in order to maintain fitness and activity levels compared to subordinates in their groups. The energetic costs of defending territory, mates, and other resources can be very consuming and cause high-ranking individuals, who spend more time in these activities, to lose body mass over long periods of dominance. Therefore, their physical condition decreases the longer they spend partaking in these high-energy activities, and they lose rank as a function of age.[2]
In wild male baboons, the highest ranking male, also known as the alpha, experiences high levels of both testosterone and glucocorticoid, which indicates that high-ranking males undergo higher levels of stress which reduces fitness. Reduced health and longevity occurs because these two hormones have immunosuppressant activity, which reduces survival and presents opportunities for parasitic infestation and other health risks. This reduced fitness due to the alpha position results in individuals maintaining high rank for shorter periods of time and having an overall reduced health and longevity from the physical strain and costs of the position.[10]
Subordinance
Benefits
There are a number of benefits to being subordinate. Subordination is beneficial in agonistic conflicts where rank predicts the outcome of a fight. Less injury will occur if subordinate individuals avoid fighting with higher-ranking individuals who would win a large percentage of the time - knowledge of the pecking order keeps both parties from incurring the costs of a prolonged fight. In hens it has been observed that both dominants and subordinates benefit from a stable hierarchical environment because fewer challenges means more resources can be dedicated to laying eggs.[11] It is also possible that in groups consisting of highly related individuals, kin selection influences the stability of hierarchical dominance. If a subordinate individual is closely related to the dominant individual, the subordinate may benefit more genetically by assisting the dominant individual, as his or her genes are still passed along in the offspring of the dominant individual or his or her genes have been passed along after possessing rank for some time. In a study of male savanna baboons, alpha males exhibited high levels of testosterone and stress. This stress and testosterone over a long period of time can lead to decreased fitness. The lowest ranking males in the hierarchy also demonstrated high stress levels, suggesting that it is the beta males that gain the most fitness, avoiding stress while still maintaining some of reproductive and nutritional benefits of moderate rank.[10]
Savanna baboons display a number of mating tactics correlated with their age. One such tactic attributed to older, subordinate males involves forming alliances to combat higher-ranking males in order to achieve access to females for copulation. These lowest ranking males would get no opportunity to copulate otherwise.[12]
Fighting with the dominant male(s) is a risky behavior that may result in defeat, injury or even death. In bighorn sheep, however, subordinates occasionally win a fight for a female, and they father 44% of the lambs born in the population. These sheep live in large flocks, and dominance hierarchies are often restructured each breeding season.[13]
Burying beetles, which have a social order involving one dominant male controlling most access to mates, display a behavior known as sneak copulation. While one male at a carcass has a 5:1 mating advantage, subordinate males will tempt females away from the carcass with pheromones and attempt to copulate, before the dominant male can drive them forcefully away.[14] In flat lizards, young males take advantage of their underdeveloped secondary sex characteristics to engage in sneak copulations. These young males mimic all the visual signs of a female lizard in order to successfully approach a female and copulate without detection by the dominant male. This strategy does not work at close range because the chemical signals given off by the sneaky males reveal their true nature, and they are chased out by the dominant.[15]
Costs
Decreased fitness and reduced access to nutrition
Subordinate individuals suffer a range of costs from dominance hierarchies, one of the most notable being reduced access to food sources. When a resource is obtained dominant individuals are first to feed as well as taking the longest time. Subordinates also lose out in shelter and nesting sites. Brown hyenas, which display defined linear dominance in both sexes, allow subordinate males and females decreased time of feeding at a carcass.[16] In toque monkeys subordinates are often displaced from feeding sites by dominant males. Additionally, they are excluded from sleeping sites, and they suffer reduced growth and increased mortality.[17]
Decreased reproductive success
Subordinate individuals often demonstrate a huge reproductive disadvantage in dominance hierarchies. Among brown hyenas, subordinate females have less opportunity to rear young in the communal den, and thus had decreased survival of offspring when compared to high-ranking individuals. Subordinate males have far less copulations with females compared to the high-ranking males.[16] In African wild dogs which live in social packs separated into male and female hierarchies, top ranking alpha females have been observed to produce 76-81% of all litters.[18]
Mitigating the costs
Subordinate animals engage in a number of behaviors in order to outweigh the costs of low rank. Dispersal is often associated with increased mortality and subordination may decrease the potential benefits of leaving the group. In the red fox it has been shown that subordinate individuals, given the opportunity to desert, often do not due to the risk of death and the low possibility that they would establish themselves as dominant members in a new group.[19] There is also the possibility that a subordinate individual who stays may become a high-ranking individual at a future time if the alpha male dies or is usurped.
Conflict over dominance
Patterns of animal conflict reveal important insights into the evolution of behavior and the influence of behavior on relationships that develop in a social group. Pair-wise interactions have been observed to promote social hierarchies within groups of animals where individuals with successful agonistic behaviors often achieve dominance. These behaviors, which include aggression, threat displays, and fighting, are indicative of competition over resources, such as food or mates. However, they may vary based on the situation and position of the individuals involved.
Animal decisions regarding involvement in conflict are defined by the interplay between the costs and benefits of agonistic behaviors. When initially developed, game theory, the study of optimal strategies during pair-wise conflict, was grounded in the false assumption that animals engaged in conflict were of equal fighting ability. Modifications, however, have provided increased focus on the differences between the fighting capabilities of animals and raised questions about their evolutionary development. These differences are believed to determine the outcomes of fights, their intensity, and animal decisions to submit or continue fighting. The influence of aggression, threats, and fighting on the strategies of individuals engaged in conflict has proven integral to establishing social hierarchies reflective of dominant-subordinate interactions.[1]
The asymmetries between individuals have been categorized into three types of interactions.[20]
- Resource-holding potential: Animals that are better able to defend resources often win without much physical contact.
- Resource value: Animals more invested in a resource are likely to invest more in the fight despite potential for incurring higher costs.
- Intruder retreats: When participants are of equal fighting ability and competing for a certain territory, the resident of the territory is likely to end as the victor because he values the territory more. This can be explained further by looking at the example of the common shrews. If one participant believes he is the resident of the territory, he will win when the opponent is weaker or food is scarce. However, if both shrews believe they are the true territory holder, the one with the greater need for food, and therefore, the one that values the resource more, is most likely to win.
As expected, the individual who emerges triumphant is rewarded with the dominant status, having demonstrated his/her physical superiority. However, the costs incurred to the defeated, which include loss of reproductive opportunities and quality food, can hinder the individual's fitness. In order to minimize these losses, animals generally retreat from fighting or displaying fighting ability unless there are obvious cues indicating victory. These often involve characteristics that provide an advantage during agonistic behavior, such as size of body, displays, etc. Red stags, for example, engage in exhausting roaring contests to exhibit their strength (Huntingford). However, such an activity would impose more costs than benefits for unfit stags, and compel them to retreat from the contest. Larger stags have also been known to make lower-frequency threat signals, acting as indicators of body size, strength, and dominance.
Engaging in agonistic behavior can be very costly and thus there are many examples in nature of animals who achieve dominance in more passive ways. In some, the dominance status of an individual is clearly visible, eliminating the need for agonistic behavior. In wintering bird flocks, white-crowned sparrows display a unique white plumage; the higher the percentage of the crown that consists of white feathers, the higher the status of the individual.[21] For other animals, the time spent in the group serves as a determinant of dominance status. Pack members of gray wolves, for example, need the time to reach the top of the ladder. Rank may also be acquired from maternal dominance rank. In rhesus monkeys, offspring gain dominance status based on the rank of the mother—the higher ranked the mother, the higher ranked the offspring will be (Yahner). Similarly, the status of a male Canada goose is determined by the rank of his family. Although dominance is determined differently in each case, it is influenced by the relationships between members of social groups.[22]
These observed interactions reflect the battle for resources. The association between resource availability and agonistic behavior suggests that animal conflict is adaptive by enabling competition and exploitation of available food and mates. The outcome of these interactions results in important social patterns that define hierarchies and, therefore, future access to resources.
Regulation mechanisms
The most basic interaction that establishes a Dominance Hierarchy is the dyad, or paired interaction among individuals. To study the formation of hierarchies, scientists have often used the dyadic method, in which two individuals are forced to interact isolated from others. All individuals in the group are paired with each other (i.e. a round-robin), in isolation, until a hierarchy can be deduced. The process of deducing the hierarchy involves the construction of a dominance matrix, in which wins/ties are expressed in relation to each member of the group.
Individuals with greater hierarchical status tend to displace those ranked lower from access to space, to food and to mating opportunities. Thus, individuals with higher social status tend to have greater reproductive success by mating more often and having more resources to invest in the survival of offspring. Hence, hierarchy serves as an intrinsic factor for population control, ensuring adequate resources for the dominant individuals and thus preventing widespread starvation. Territorial behavior enhances this effect.[23][24]
These hierarchies are not fixed and depend on any number of changing factors, among them are age, gender, body size, intelligence, and aggressiveness. For instance, in linear hierarchies, a lost top ranked individual ("alpha") is usually replaced by its direct subordinate ("beta"); this individual assumes its role in the group and gains the same benefits. In eusocial species, decrease of fertility is among the main reasons for ranking displacement.
In eusocial organisms
The suppression of reproduction by dominant individuals is the most common mechanism that maintains the hierarchy. In eusocial mammals this is mainly achieved by aggressive interactions between the potential reproductive females. In eusocial insects, aggressive interactions between sexuals are common determinants of reproductive status, such as in the bumblebee Bombus bifarius,[25] the paper wasp Polistes annularis[26] and in the ants Dinoponera australis and D. quadriceps.[27] In general aggressive interactions are ritualistic and involve antennation (drumming), abdomen curling and very rarely mandible bouts and stinging. The winner of the interaction may walk over the subordinated, that in turn assumes a prostrated posture on the substrate. In order to be effective, these regulatory mechanisms must include traits that make an individual rank position readily recognizable by its nestmates. The composition of the lipid layer present on the cuticle of social insects is the clue used by nestmates to recognize each other in the colony, and to discover each insect's reproductive status (and therefore its rank).[28] Visual cues may also transmit the same information. Paper wasps Polistes dominulus have individual "facial badges" that permit them to recognize each other and to identify the status of each individual.[29] Individuals whose badges were modified by painting were aggressively treated by their nestmates, suggesting that advertising a false ranking status is a costly behavior, suppressed in these wasps.
Other behaviors are involved in maintaining reproductive status in social insects. The removal of a thoracic sclerite in Diacamma ants inhibits ovary development; the only reproductive individual of this naturally queenless genus is the one that retains its sclerite intact. This individual is called a gamergate, and is responsible for mutilating all the newly emerged females, to maintain its social status. Gamergates of Harpegnathos saltator arise from aggressive interactions, forming a hierarchy of potential reproductives.[30]
In the honey bee Apis mellifera, pheromone produced by the queen mandibular glands is responsible for inhibiting ovary development in the worker caste.[31] "Worker policing" is an additional mechanism that prevents reproduction by workers, found in bees and ants. Policing may involve oophagy and immobilization of egg-layers from the worker caste.[32] In some ant species such as the carpenter ant Camponotus floridanus, eggs from queens have a peculiar chemical profile that workers can distinguish from worker laid eggs. When worker-laid eggs are found, they are eaten.[33] In some species, such as Pachycondyla obscuricornis, workers may try to escape policing by shuffling their eggs within the egg pile laid by the queen.[34]
Hormones and dominance
It has been shown that modulation of hormone levels after hibernation may be associated with the establishment of dominance hierarchies within the social order of the paper wasp (Polistes dominulus).[35] Former studies have shown that dominance hierarchies within the paper wasp species is dependent on the queen (also known as a foundress), though the mechanism behind the establishment is currently under investigation. Recent research suggests that this manifestation may be dependent on specific hormones contained within the foundress. Laboratory experiments have shown that when foundresses are injected with juvenile hormone (JH), a hormone responsible for regulating growth and development in many insect species including wasps, the foundresses exhibit an increase in dominant behavior compared to those who had not been injected.[35] Further evidence shows that foundresses that have a larger corpora allata, a region of the female wasp brain responsible for the synthesis and secretion of JH, are naturally more dominant.[35] In an effort to determine if JH is indeed responsible for manifesting dominance, a follow up experiment was done utilizing 20-hydroxyecdysone, an ecdysone known to enhance maturation and size of oocytes.[35] The size of the oocytes plays a significant role in establishing dominance within Polistes dominulus and thus researchers used this hormone to compare which condition (either JH or 20-hydroxyecdysone treated) induced higher levels of dominance intensities within foundresses.[36] The results showed that the 20-hydroxyecdysone treated foundresses showed increased dominance compared to those foundresses treated with JH and suggests that 20-hydroxyecdysone, not JH may play a larger role in establishing dominance (Roseler et al., 1984). Subsequent research however, suggests that JH is implicated in the manifestation of dominance, though it only exerts dominance-type effects on certain individuals. By recording the number of mounting attempts between rival foundresses as a measure of dominance, researchers found that when injected with the same amount of JH, larger foundresses showed more mounting behaviors compared to foundresses that were smaller. Additionally, the more dominant foundress tended to show an increased number of oocytes contained within her ovaries. It was concluded that larger foundresses are more reproductively fit and thus JH, which is responsible for the growth and maturation of the ovaries, be more active within these individuals compared to the smaller, less fertile foundresses though the mechanism of action and/or any synergistic effects between JH and other hormones remains unknown.[36]
The hormone model of dominance and reproductive capacity has also been demonstrated in the naked mole-rat (Heterocephalus glaber). It has previously been established that the dominance hierarchy within the species is dependent on the highest ranking female (queen) and her ability to suppress critically important reproductive hormones in male and female sub-dominants. In sub-dominant males, appears that lutenizing hormone and testosterone are suppressed while in females it appears that the suppression involves the entire suppression of the ovarian cycle. This suppression reduces sexual virility and behavior and thus redirects the sub-dominant's behavior into helping the queen with her offspring.[37] though the mechanisms of how this is accomplished are debated. Former research suggests that primer pheromones secreted by the queen cause direct suppression of these vital reproductive hormones and functions however current evidence suggests that it is not the secretion of pheromones which act to suppress reproductive function but rather the queen's extremely high levels of circulating testosterone, which cause her to exert intense dominance and aggressiveness on the colony and thus "scare" the other mole-rats into submission.[38] Research has shown that removal of the queen from the colony allows the reestablishment of reproductive function in sub-dominant individuals. To see if a priming pheromone secreted by the queen was indeed causing reproductive suppression, researchers removed the queen from the colony but did not remove her bedding. They reasoned that if a primer pheromones were on the bedding then the sub-dominant's reproductive function should continue to be suppressed. Instead however, they found that the sub-dominants quickly regained reproductive function even in the presence of the queen's bedding and thus it was concluded that primer pheromones do not seem to play a role in suppressing reproductive function.[38]
Evidence also suggests that glucocorticoids, signaling molecules (commonly known as stress hormones) produced by the adrenal glands which stimulate the fight or flight response may also be implicated in the establishment of dominance hierarchies. Field research has shown that higher ranking individuals tend to have much higher levels of circulating glucocorticoids compared to subdominant individuals within the hierarchy.[39] This is in stark opposition to the original suggestion stating that being subdominant was more stressful than being dominant within a hierarchy.[40] Two core hypotheses attempt to explain these unusual findings. The first suggests that higher ranking individuals exert more energy and thus need higher levels of glucocorticoids to more readily mobilize glycogen stores for energy use.[41] This hypothesis is supported by research showing that when food availability is low, cortisol levels tended to increase within the dominant male thus allowing increased glycogen metabolism and subsequent energy production.[40] The second hypothesis suggests that elevated stress hormones are a result of social factors within the hierarchy particularly when the hierarchy is in transition. Proponents of this theory assert that when a hierarchy is unstable, aggressive interaction and confrontations increase within the hierarchy. As a result, the dominant individual ends up fighting much more than when the hierarchy is stable and as consequence of the increased fighting, glucocorticoids are elevated during this period. Field studies of olive baboons in Kenya seem to support this hypothesis as it has been observed that dominant individuals tended to have lower cortisol levels in a stable hierarchy than did subdominant individuals. When the hierarchy shifted to being unstable however, the dominant individual showed much higher levels of cortisol compared to subdominant individuals.[42] Many are quick to point out however, that this is extremely unusual as dominant individuals rarely have low levels of glucocorticoids regardless of hierarchy state.[40] Nonetheless, it appears that being dominant may not be as beneficial as previously thought since chronic levels of glucocorticoids can have a variety of adverse health effects including immunosuppression, increased appetite, and osteoporosis (see Cortisol) and is a possible reason why subdominant individuals are reluctant to challenge the dominant individual for their position during hierarchy transitory periods.
Brain pathways and hierarchy
Several areas of the brain contribute to hierarchical behavior in animals. One of the areas that has been linked with this behavior is the prefrontal cortex, a region involved with decision making and social behavior. High social rank in a hierarchical group of mice has been associated with increased excitability in the medial prefrontal cortex of pyramidal neurons, the primary excitatory cell type of the brain.[43] High ranking macaques have a larger rostral prefrontal cortex in large social groups.[44] Additionally, neuroimaging studies with computer stimulated hierarchal conditions, led to increased activity in the ventral and dorsolateral prefrontal cortex, one processing judgment cues and the other processing status of an individual. Other studies have determined that lesions to the prefrontal cortex (when the area is severed to disrupt functioning to observe its role in behavior) led to deficits in processing social hierarchy cues, suggesting this area is important in regulating this information.[45] Although the prefrontal cortex has been implicated, there are other downstream targets of the prefrontal cortex that have also been linked in maintaining this behavior. This includes the amygdala through lesion studies in rats and primates which led to disruption in hierarchy, and can affect the individual negatively or positively depending on the subnuclei that is targeted. Additionally, the dorsal medial PFC-medial dorsal thalamus connection has been linked with maintenance of rank in mice.[46] Another area that has been associated is the dorsal raphe nucleus, the primary serotonergic nuclei (a neurotransmitter involved with many behaviors including reward and learning). In manipulations studies of this region, there were changes in fighting and affiliative behavior in primates and crustaceans.[45]
Interpersonal complementarity hypothesis
The interpersonal complementarity hypothesis suggests that obedience and authority are reciprocal, complementary processes. That is, it predicts that one group member's behaviours will elicit a predictable set of actions from other group members. Friendly behaviours are predicted to be met with friendly behaviours, and hostile behaviours are predicted to be reciprocated with similar, hostile behaviours. When an individual acts in a dominant, authoritative manner in a group, this behaviour tends to prompt submissive responses from other group members. Similarly, when group members display submissive behaviour, others feel inclined to display dominant behaviours in return. Tiedens and Fragle (2003) found that hierarchical differentiation plays a significant role in liking behaviour in groups.[47] Individuals prefer to interact with other group members whose power, or status behaviour complements their own. That is to say, group members who behave submissively when talking to someone who appears to be in control are better liked, and similarly individuals who display dominant behaviours (e.g., taking charge, issuing orders) are more liked when interacting with docile, subservient individuals.
In specific biological classes
Female dominance in mammals
Female-biased dominance occurs rarely in mammals. It occurs when all adult males exhibit submissive behavior to adult females in social settings. These social settings are usually related to feeding, grooming, and sleeping site priority. It is observed consistently in hyenas, lemurs and the bonobo.[48] It is also observed in elephants. The ring-tailed lemur is observed to be the most prominent model of female dominance.[49]
There are three basic proposals for the evolution of female dominance:[50]
- The Energy Conservation Hypothesis: males subordinate to females to conserve energy for intense male-male competition experienced during very short breeding seasons
- Female behavioral strategy: dominance helps females deal with the unusually high reproductive demands; they prevail in more social conflicts because they have more at stake in terms of fitness.
- Male behavioral strategy: males defer as a parental investment because it ensures more resources in a harsh unpredictable climate for the female, and thus, the male's future offspring.
Since these original proposals, scientists like Peter Kappeler have modified and integrated other ideas.
In the case of lemurs, there is no single hypothesis that can fully explain female social dominance at this time and all three are likely to play a role. Adult female lemurs have increased concentrations of angrogens when they transition from non-breeding to breeding seasons, increasing female aggression.[51] In ring-tailed lemurs, androgens are greater in pregnant females, which suggests that organizational androgens might influence the developing offspring.[52] Organizational androgens play a role in "explaining female social dominance" in ring-tailed lemurs, as androgens are associated with aggressive behavior in young females.[53] Females that were "exposed to greater concentrations of maternal [androtenedione] late in fetal development were less likely to be aggressed against postnatally, whereas females that were...exposed to greater concentrations of maternal [testosterone]...were more likely to receive aggression postnatally."[53]
Dominance rank in female chimpanzees correlates positively with reproductive success. Although a high rank obviously has an advantage for females, clear (linear) hierarchies in female chimpanzees have not been detected.[54]
In "masculinized" female mammals like the spotted hyena (Crocuta crocuta), androgens (i.e. specifically, androstenedione and testosterone) are "implicated in the organization and activation of...nonreproductive behavioral traits, including aggression, social dominance, rough-and-tumble play, and scent marking"[55] For aggressively dominant female meerkats (Suricata suricatta), they have "exceptionally high concentrations" of androgens, "particularly during gestation"[56]
Birds
In dunnocks, two or three males may share one or more females in a polyandrous or polygynandrous mating system. This leads to the males' ranges to overlap so they would cooperate in order to defend the territory. Among the males is a dominance hierarchy: older birds tend to be the alpha male and first year birds are usually the beta males. Studies have not found cases of close male relatives being involved in any mating combinations.
The blue-footed booby brood of two chicks always has a dominance hierarchy due to the asynchronous hatching of the eggs. One egg is laid four days before the other, and incubation starts immediately after laying, so the elder chick is hatched four days before the younger chick and has a four-day head start on growth. The elder, stronger chick almost always becomes the dominant chick. During times of food shortage, the dominant chick often kills the subordinate chick by either repeatedly pecking or by ousting the younger chick from the nest. The brood hierarchy makes it easier for the subordinate chick to die quietly in times of food scarcity, which provides an efficient system for booby parents to maximize their investment.[57]
Fish
Dominance hierarchies can also be observed in many fish. For example, in the dragonet, males form hierarchies that are often exhibited during mating. They can act extremely aggressively towards another male if it intrudes upon courtship and pairing with a female, and fights can be very intense.
Eusocial insects
In insect societies, only one to few individuals members of a colony can reproduce, whereas the other colony members have their reproductive capabilities suppressed. This conflict over reproduction in some cases results in a dominance hierarchy. Dominant individuals in this case are known as queens and have the obvious advantage of performing reproduction and benefiting from all the tasks performed by their subordinates, the worker caste (foraging, nest maintenance, nest defense, brood care and thermal regulation). According to Hamilton's rule, the reproduction costs of the worker caste are compensated by the contribution of workers to the queen's reproductive success, with which they share genes. This is true not only to the popular social insects (ants, termites, some bees and wasps), but also for the naked mole-rat Heterocephalus glaber. In a laboratory experiment, Clarke and Faulkes (1997)[58] demonstrated that reproductive status in a colony of H. glaber was correlated with the individual's ranking position within a dominance hierarchy, but aggression between potential reproductives only started after the queen was removed.
Social insects above mentioned, excluding termites, are haplodiploid. Queen and workers are diploid, but males develop from haploid genotypes. In some species, suppression of ovary development is not totally achieved in the worker caste, which opens the possibility of reproduction by workers. Since nuptial flights are seasonal and workers are wingless, workers are almost always virgin, and (as gamergate ants or laying worker bees) can only lay unfertilised eggs. These eggs are in general viable, developing into males. A worker that perform reproduction is considered a 'cheater' within the colony, because its success in leaving descendants becomes disproportionally larger, compared to its sisters and mother. The advantage of remaining functionally sterile is only accomplished if every worker assume this 'compromise'. When one or more workers start reproducing, the 'social contract' is destroyed and the colony cohesion is dissolved. Aggressive behavior derived from this conflict may result in the formation of hierarchies, and attempts of reproduction by workers are actively suppressed. In some wasps, such as Polistes fuscatus, instead of not laying eggs, the female workers begin being able to reproduce, but once being under the presence of dominant females, the subordinate female workers can no longer reproduce.[59]
In some wasp species such as Liostenogaster flavolineata there are many possible Queens that inhabit a nest, but only one can be queen at a time. When a Queen dies the next Queen is selected by an age based dominance hierarchy. This is also true in the species Polistes instabilis, where the next queen is selected based on age rather than size. Polistes exclamans also exhibits this type of hierarchy.[60] Within the dominance hierarchies of the Polistes versicolor, however, the dominant-subordinate context in the yellow paper wasps is directly related to the exchange of food. Future foundresses within the nest compete over the shared resources of nourishment, such as protein. Unequal nourishment is often what leads to the size differences that result in dominant-subordinate position rankings. Therefore, if during the winter aggregate, the female is able to obtain greater access to food, the female could thus reach a dominant position.[61]
In some species, especially in ants, more than one queen can be found in the same colony, a condition called Polygyny. In this case, another advantage of maintaining a hierarchy is to prolong the colony lifespan. The top ranked individuals may die or lose fertility and "extra queens" may benefit of starting a colony in the same site or nest. This advantage is critical in some ecological contexts, such as in situations where nesting sites are limited or dispersal of individuals is risky due to high rates of predation. This polygynous behavior has also been observed in some eusocial bees such as Schwarziana quadripunctata. In this species, multiple queens of varying sizes are present. The larger, physogastric, queens typically control the nest, though a "dwarf" queen will take its place in the case of a premature death.[62]
Variations
Egalitarian and despotic social systems
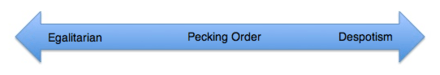
The manifestation of intrasexual conflict can be observed in one of two systems. The social order can be either egalitarian or despotic. In a linear ranking system (often referred to as a pecking order), every member of the gender is recognized as either dominant or submissive relative to every other member, creating a linear distribution of rank. For example, groups of spotted hyenas and brown hyenas both demonstrate linear dominance.[63] In a despotic system, one or two members are considered dominant while all other members of the living group are equally submissive. Examples of despotic social systems are found in leopard geckos, dwarf hamsters, gorilla, Neolamprologus pulcher, and African wild dog.[64]
Reverse dominance hierarchy
Christopher Boehm, having explored data from 48 societies spread across the globe, ranging from small hunting and gathering bands to more sedentary chiefdoms, suggested that with the advent of anatomically modern humans who continued to live in small groups and had not yet domesticated plants and animals (hunter-gatherer), it is very likely that all human societies practised egalitarianism and that most of the time they did so very successfully.[65]
Boehm writes:
- "As long as followers remain vigilantly egalitarian because they understand the nature of domination and leaders remain cognizant of this ambivalence-based vigilance, deliberate control of leaders may remain for the most part highly routinized and ethnographically unobvious."
Boehm identifies the following mechanisms ensuring the Reverse Dominance Hierarchy: Public Opinion, Criticism and Ridicule, Disobedience, and Extreme Sanctions. Further characteristics include ambivalence towards leaders and anticipation of domination.[65]
See also
- Deference
- Evolution of intelligence
- Hierarchical organization
- Hypergamy
- Pecking order
- Rankism
- Social class
- Social organization
- Sociobiology
References
- Chase, I. D.; Tovey, C.; Murch, P. (2003). "Two's Company, Three's a Crowd: Differences in Dominance Relationships in Isolated versus Socially Embedded Pairs of Fish". Behaviour. 140 (10): 1193–217. doi:10.1163/156853903771980558. S2CID 56345496.
- Huntingford, Felicity, and Angela K. Turner. Animal Conflict. London: Chapman and Hall, 1987
- Cowlishaw, Guy; Dunbar, Robin IM (1991). "Dominance rank and mating success in male primates". Animal Behaviour. 41 (6): 1045–1056. doi:10.1016/s0003-3472(05)80642-6.
- Von Rueden, Christopher R.; Jaeggi, Adrian V. (2016). "Men's status and reproductive success in 33 nonindustrial societies: Effects of subsistence, marriage system, and reproductive strategy". Proceedings of the National Academy of Sciences. 113 (39): 10824–10829. doi:10.1073/pnas.1606800113. PMC 5047206. PMID 27601650.
- Samuels, A.; Silk, J. B.; Rodman, P. (1984). "Changes in the dominance rank and reproductive behavior of male bonnet macaques (Macaca radiate)". Anim. Behav. 32 (4): 994–1003. doi:10.1016/s0003-3472(84)80212-2.
- Widdig, Anja; Kessler, Matthew J.; Bercovitch, Fred B.; Berard, John D.; Duggleby, Christine; Nürnberg, Peter; Rawlins, Richard G.; et al. (2016). "Genetic studies on the Cayo Santiago rhesus macaques: a review of 40 years of research". American Journal of Primatology. 78 (1): 44–62. doi:10.1002/ajp.22424. PMID 26031601.
- Dubuc, Constance; Muniz, Laura; Heistermann, Michael; Engelhardt, Antje; Widdig, Anja (2011). "Testing the priority-of-access model in a seasonally breeding primate species". Behavioral Ecology and Sociobiology. 65 (8): 1615–1627. doi:10.1007/s00265-011-1172-8. PMC 3134767. PMID 21874084.
- Bonnatti, Vanessa; Luz Paulino Simões, Zilá; Franco, Fernando Faria; Tiago, Mauricio (3 January 2014). "Evidence of at least two evolutionary lineages in Melipona subnitida (Apidae, Meliponini) suggested by mtDNA variability and geometric morphometrics of forewings". Naturwissenschaften. 101 (1): 17–24. Bibcode:2014NW....101...17B. doi:10.1007/s00114-013-1123-5. PMID 24384774.
- Koedam, D.; Contrera, A. de O. Fidalgo; Imperatriz-Fonseca, V. L. (2004). "How queen and workers share in male production in the stingless bee Melipona subnitida Ducke (Apidae, Meliponini)". Insectes Sociaux. 52 (2): 114–121. doi:10.1007/s00040-004-0781-x.
- Gesquiere, Laurence R.; Learn, Niki H.; Simao, Carolina M.; Onyango, Patrick O.; Alberts, Susan C.; Altmann, Jeanne (2011). "Life at the Top: Rank and Stress in Wild Male Baboons". Science. 333 (6040): 357–60. Bibcode:2011Sci...333..357G. doi:10.1126/science.1207120. PMC 3433837. PMID 21764751.
- Pusey, A.E., and C. Packer. 1997. The ecology of relationships. In Behavioural Ecology: An Evolutionary Approach, edited by J. R. Krebs and N.B. Davies. Oxford: Blackwell Science, pp 254–283
- Noe, R.; Sluijter, A. A. (1990). "Reproductive Tactics of Male Savanna Baboons". Behaviour. 113 (1–2): 117–170. doi:10.1163/156853990x00455. S2CID 14258881.
- Hogg, J. T.; Forbes, S. H. (1997). "Mating in bighorn sheep: Frequent male reproduction via a high-risk unconventional tactic". Behavioral Ecology and Sociobiology. 41 (1): 33–48. doi:10.1007/s002650050361.
- Pettinger, Adam M.; Steiger, Sandra; Mueller, Josef K.; Sakaluk, Scott K.; Eggert, Anne-Katrin (2011). "Dominance status and carcass availability affect the outcome of sperm competition in burying beetles". Behavioral Ecology. 22 (5): 1079–1087. doi:10.1093/beheco/arr093.
- Whiting, Martin J.; Webb, Jonathan K.; Keogh, J. Scott (2009). "Flat lizard female mimics use sexual deception in visual but not chemical signals". Proceedings of the Royal Society B: Biological Sciences. 276 (1662): 1585–1591. doi:10.1098/rspb.2008.1822. PMC 2660994. PMID 19324828.
- Owens, D.; Owens, M. (1996). "Social dominance and reproductive patterns in brown hyaenas, Hyaena brunnea, of the central Kalahari desert". Animal Behaviour. 51 (3): 535–551. doi:10.1006/anbe.1996.0058.
- Dittus, W. P. J. (1977). "The Social Regulation of Population Density and Age-Sex Distribution in the Toque Monkey". Behaviour. 63 (3): 281–322. doi:10.1163/156853977x00450.
- Creel, S. (1997). "Handling of African wild dogs and chronic stress: Reply". Conservation Biology. 11 (6): 1454–1456. doi:10.1046/j.1523-1739.1997.0110061454.x.
- Baker, P. J.; Robertson, C. P. J.; Funk, S. M.; Harris, S. (1998). "Potential fitness benefits of group living in the red fox, Vulpes vulpes". Animal Behaviour. 56 (6): 1411–1424. doi:10.1006/anbe.1998.0950. PMID 9933538. S2CID 24702890.
- Huntingford, Felicity. The Study of Animal Behaviour. London: Chapman and Hall, 1984.
- Laubach, Zachary. "Functional Importance of Plumage Badges as Intraspecific Signals in White-Crowned Sparrows (zonotrichia Leucophrys Oriantha) : Deep Blue at the University of Michigan." Functional Importance of Plumage Badges as Intraspecific Signals in White-Crowned Sparrows (zonotrichia Leucophrys Oriantha) : Deep Blue at the University of Michigan. Deep Blue. 27 Nov. 2012. <http://hdl.handle.net/2027.42/77948>.
- Yahner, Richard H. Wildlife Behavior and Conservation. New York: Springer, 2012.
- "Behavior: The Animal Watchers". Time. 1973-10-22. Retrieved 2010-05-01.
- Lorenz:On Aggression:BOOK SUMMARY
- Foster, R. L.; Ameilia, B.; Verdirame, D.; O'Donnell, S (2004). "Reproductive physiology, dominance interactions, and division of labour among bumble bee workers". Physiological Entomology. 29 (4): 327–334. doi:10.1111/j.0307-6962.2004.00388.x.
- Hughes, CR; Beck, MO; Strassman, J. E. (1987). "Queen succession in the social wasp Polistes annularis". Ethology. 76 (2): 124–132. doi:10.1111/j.1439-0310.1987.tb00678.x.
- Monnin, T.; Ratnieks, F. L. W.; Brandao, C. R. F. (2003). "Reproductive conflict in animal societies: hierarchy length increases with colony size in queenless ponerine ants". Behavioral Ecology and Sociobiology. 54: 71–79. doi:10.1007/s00265-003-0600-9.
- Monnin, T. (2006). "Chemical recognition of reproductive status in social insects". Annales Zoologici Fennici. 43: 515–530.
- Tibbetts, E. A.; Dale, J. (2004). "A socially enforced signal of quality in paper wasp". Nature. 432 (7014): 218–222. Bibcode:2004Natur.432..218T. doi:10.1038/nature02949. PMID 15538369.
- Peeters, C.; Liebig, J.; Hölldobler, B. (2000). "Sexual reproduction by both queens and workers in the ponerine ant Harpegnathos saltator". Insectes Sociaux. 47 (4): 325–332. doi:10.1007/pl00001724.
- Hoover, S. E. R.; Keeling, C. I.; Winston, M. L.; Slessor, K. N. (2003). "The effect of queen pheromones on worker honey bee ovary development". Naturwissenschaften. 90 (10): 477–480. Bibcode:2003NW.....90..477H. doi:10.1007/s00114-003-0462-z. PMID 14564409.
- Ratnieks, FLW; Visscher, PK. (1989). "Worker policing in the honeybee". Nature. 342 (6251): 796–797. Bibcode:1989Natur.342..796R. doi:10.1038/342796a0.
- Endler, A.; Liebig, J.; Schmitt, T.; Parker, . E.; Jones, G. R.; Schreier, P; Hölldobler, B. (2004). "Surface Hydrocarbons of queen eggs regulate worker reproduction in a social insect". PNAS. 101 (9): 2945–2950. Bibcode:2004PNAS..101.2945E. doi:10.1073/pnas.0308447101. PMC 365725. PMID 14993614.
- Oliveim, PS; Hölldobler, B (1991). "Agonistic interactions and reproductive dominance in Pachycondyla obscuricornis (Hymenoptera, Formicidae)". Psyche. 98 (2–3): 215–226. doi:10.1155/1991/64635.
- Roseler, P.F.; Roseler, I.; Strambi, A.; Augier, R. (1984). "Influence of insect hormones on the establishment of dominance hierarchies among foundresses of the paper wasp, Polistes gallicus". Behavioral Ecology and Sociobiology. 15 (2): 133–142. doi:10.1007/bf00299381.
- Tibbetts, E. A.; Izzo, A.S. (2009). "Endocrine mediated phenotypic plasticity: Condition-dependent effects on juvenile hormone on dominance and fertility of wasp queens". Hormones and Behavior. 56 (5): 527–531. doi:10.1016/j.yhbeh.2009.09.003. PMID 19751736.
- Clarke, F. M.; Faulkes, C. G. (1998). Hormonal and behavioural correlates of male dominance and reproductive status in captive colonies of the naked mole-rat, Heterocephalus glaber. The Royal Society.
- Faulkes, C. G.; Abbott, D. H. (1993). "Evidence that primer pheromones do not cause social suppression of reproduction in male and female naked mole-rats". Journal of Reproduction and Fertility. 99 (1): 225–230. doi:10.1530/jrf.0.0990225. PMID 8283442.
- For example, fishes such as Oreochromis mossambicus display higher levels of glucocorticoids in more dominant individuals compared to less dominant ones. Creel, S. (2005). "Dominance, aggression and glucocorticoid levels in social carnivores". Journal of Mammalogy. 86 (2): 255–264. doi:10.1644/bhe-002.1.
- Muller, M. N.; Wrangham, R.W. (2004). "Dominance, cortisol and stress in wild chimpanzees" (PDF). Journal of Behavioral Ecology and Sociobiology. 55 (4): 332–340. doi:10.1007/s00265-003-0713-1. hdl:2027.42/46903.
- Genuth, S. M. (1993). The endocrine system. In:Berne R. M., Levy M. N. (eds) Physiology, vol 3. Mosby Year Book, St. Louis. pp 813-1024
- Sapolsky, R. M. (1992). "Cortisol concentrations and the social significance of rank instability among wild baboons". Journal of Psychoneuroendocrinology. 17 (6): 701–709. doi:10.1016/0306-4530(92)90029-7. PMID 1287688.
- Wang, F.; Zhu, J.; Zhu, H.; Zhang, Q.; Lin, Z.; Hu, H. (2011-11-04). "Bidirectional Control of Social Hierarchy by Synaptic Efficacy in Medial Prefrontal Cortex". Science. 334 (6056): 693–697. Bibcode:2011Sci...334..693W. doi:10.1126/science.1209951. ISSN 0036-8075. PMID 21960531.
- Sallet, J.; Mars, R. B.; Noonan, M. P.; Andersson, J. L.; O'Reilly, J. X.; Jbabdi, S.; Croxson, P. L.; Jenkinson, M.; Miller, K. L.; Rushworth, M. F. S. (2011-11-04). "Social Network Size Affects Neural Circuits in Macaques". Science. 334 (6056): 697–700. Bibcode:2011Sci...334..697S. doi:10.1126/science.1210027. ISSN 0036-8075. PMID 22053054.
- Kessels, Helmut W Hu, Hailan Wang, Fei (2014). The mouse that roared: neural mechanisms of social hierarchy. OCLC 931061361.CS1 maint: multiple names: authors list (link)
- Zhou, Tingting; Zhu, Hong; Fan, Zhengxiao; Wang, Fei; Chen, Yang; Liang, Hexing; Yang, Zhongfei; Zhang, Lu; Lin, Longnian; Zhan, Yang; Wang, Zheng (2017-07-14). "History of winning remodels thalamo-PFC circuit to reinforce social dominance". Science. 357 (6347): 162–168. Bibcode:2017Sci...357..162Z. doi:10.1126/science.aak9726. ISSN 0036-8075. PMID 28706064.
- Tiedens, Larissa; Fragale, Alison (2003). "Power moves: Complementarity in dominant and submissive nonverbal behavior". Journal of Personality and Social Psychology. 84 (3): 558–568. CiteSeerX 10.1.1.319.709. doi:10.1037/0022-3514.84.3.558.
- Digby, L. I.; Kahlenberg, S. M. (2002). "Female dominance in blue-eyed black lemurs". Primates. 43 (3): 191–199. doi:10.1007/BF02629647. PMID 12145400.
- Sauther, Michelle L. (1993), Lemur Social Systems and Their Ecological Basis, Springer US, pp. 135–152, doi:10.1007/978-1-4899-2412-4_10, ISBN 978-1-4899-2414-8 Missing or empty
|title=(help);|chapter=ignored (help) - Young, Andrew L.; Richard, Alison F.; Aiello, Leslie C. (1990). "Female Dominance and Maternal Investment in Strepsirhine Primates". The American Naturalist. 135 (4): 473–488. doi:10.1086/285057.
- Drea, Christine M. (April 2007). "Sex and seasonal differences in aggression and steroid secretion in Lemur catta: Are socially dominant females hormonally 'masculinized'?". Hormones and Behavior. 51 (4): 555–567. doi:10.1016/j.yhbeh.2007.02.006. ISSN 0018-506X. PMID 17382329.
- Drea, Christine M. (August 2009). "Endocrine Mediators of Masculinization in Female Mammals". Current Directions in Psychological Science. 18 (4): 221–226. doi:10.1111/j.1467-8721.2009.01640.x. ISSN 0963-7214.
- Grebe, Nicholas M.; Fitzpatrick, Courtney; Sharrock, Katherine; Starling, Anne; Drea, Christine M. (September 2019). "Organizational and activational androgens, lemur social play, and the ontogeny of female dominance". Hormones and Behavior. 115: 104554. doi:10.1016/j.yhbeh.2019.07.002. ISSN 0018-506X. PMID 31276664.
- Wittig, Roman M.; Boesch, Christophe (2003-08-01). "Food Competition and Linear Dominance Hierarchy Among Female Chimpanzees of the Taï National Park". International Journal of Primatology. 24 (4): 847–867. doi:10.1023/A:1024632923180. ISSN 1573-8604.
- Drea, C. M.; Weldele, M. L.; Forger, N. G.; Coscia, E. M.; Frank, L. G.; Licht, P.; Glickman, S. E. (1998-05-01). "Androgens and masculinization of genitalia in the spotted hyaena (Crocuta crocuta). 2. Effects of prenatal anti-androgens". Reproduction. 113 (1): 117–127. doi:10.1530/jrf.0.1130117. ISSN 1470-1626. PMID 9713384.
- Davies, Charli S.; Smyth, Kendra N.; Greene, Lydia K.; Walsh, Debbie A.; Mitchell, Jessica; Clutton-Brock, Tim; Drea, Christine M. (December 2016). "Exceptional endocrine profiles characterise the meerkat: sex, status, and reproductive patterns". Scientific Reports. 6 (1): 35492. doi:10.1038/srep35492. ISSN 2045-2322. PMC 5067592. PMID 27752129.
- Drummond, Hugh; Edda Gonzalez; Jose Luis Osorno (1986). "Parent-Offspring Cooperation in the Blue-footed Booby (Sula nebouxii): Social Roles in Infanticidal Brood Reduction". Behavioral Ecology and Sociobiology. 19 (5): 365–372. doi:10.1007/bf00295710.
- Clarke, F. M.; Faulkes, C. G. (1997). "Dominance and queen succession in captive colonies of the eusocial naked mole-rat, Heterocephalus glaber". Proc Biol Sci. 264 (1384): 993–1000. Bibcode:1997RSPSB.264..993C. doi:10.1098/rspb.1997.0137. PMC 1688532. PMID 9263466.
- West-Eberhard, M. J. (1969). "The social biology of polistine wasps". Mis. Publ. Zool. Univ. Michigan. 140: 1–101.
- Strassmann & Meyer (1983). "Gerontocracy in the social wasp, Polistes exclamans". Animal Behaviour. 31 (1): 431–438. doi:10.1016/S0003-3472(83)80063-3.
- González, J. A.; Nascimento, F. S.; Gayubo, S. F. (2002). "Observations on the Winter Aggregates of Two Polistine Paper Wasps (Hymenoptera Vespidae Polistinae)". Tropical Zoology. 15 (1): 1–4. doi:10.1080/03946975.2002.10531162.
- Nogueira-Ferreira, F. H.; Silva-Matos, E. V.; Zucchi, R. (2009). "Interaction and Behavior of Virgin and Physogastric Queens in Three Meliponini Species (Hymenoptera, Apidae)". Genetics and Molecular Research. 8 (2): 703–708. doi:10.4238/vol8-2kerr008. PMID 19554769.
- Watts, Heather E.; Holekamp, Kay E. (2007-08-21). "Hyena societies". Current Biology. 17 (16): R657–R660. doi:10.1016/j.cub.2007.06.002. PMID 17714659.
- Alcock, John. Animal Behavior: An Evolutionary Approach. Sunderland, MA: Sinauer Associates, 2005.
- Boehm, Christopher (1993). "Egalitarian Behavior and Reverse Dominance Hierarchy". Current Anthropology. 34 (3): 227–254. doi:10.1086/204166.
- Chase, I.; Tovey, C.; Spangler-Martin, D.; Manfredonia, M. (2002). "Individual differences versus social dynamics in the formation of animal dominance hierarchies". PNAS. 99 (9): 5744–5749. Bibcode:2002PNAS...99.5744C. doi:10.1073/pnas.082104199. PMC 122842. PMID 11960030.
- Chase, I.; Bartolomeo, C.; Dugatkin, L. (1994). "Aggressive interactions and inter-contest interval: how long do winners keep winning?". Animal Behaviour. 48 (2): 393–400. doi:10.1006/anbe.1994.1253.
- Cummins, D.D. (1996). "Dominance Hierarchies and the Evolution of Human Reasoning". Minds and Machines. 6 (4): 463–480.
- Lehner, Philip N, 1998. Handbook of ethological methods (2nd. ed.). Cambridge University Press: Cambridge, England, pp. 332–335.
- Oliveira, RF; McGregor, PK; Latruffe, C (1998). "Know thine enemy: fighting fish gather information from observing conspecific interactions". Proceedings of the Royal Society B: Biological Sciences. 265 (1401): 1045–1049. doi:10.1098/rspb.1998.0397. PMC 1689170.
- Wilson, E. O. Sociobiology. 2000.
- http://www.stanford.edu/group/stanfordbirds/text/essays/Dominance_Hierarchies.html
| Topics |
|
|---|---|
| Supporters | |
| Opponents | |
| |