Juvenile hormone
Juvenile hormones (JHs) are a group of acyclic sesquiterpenoids that regulate many aspects of insect physiology. The first discovery of a JH was by Vincent Wigglesworth. JHs regulate development, reproduction, diapause, and polyphenisms.[1][2][3]
In insects, JH (formerly called neotenin) refers to a group of hormones, which ensure growth of the larva, while preventing metamorphosis. Because of their rigid exoskeleton, insects grow in their development by successively shedding their exoskeleton (a process known as molting).
Juvenile hormones are secreted by a pair of endocrine glands behind the brain called the corpora allata. JHs are also important for the production of eggs in female insects.
JH was isolated in 1965 by Williams and Slama and the first molecular structure of a final six was solved in 1967.[4]
Most insect species contain only juvenile growth hormone (JH) III.[5] To date JH 0, JH I, and JH II have been identified only in the Lepidoptera (butterflies and moths). The form JHB3 (JH III bisepoxide) appears to be the most important JH in the Diptera, or flies.[6] Certain species of crustaceans have been shown to produce and secrete methyl farnesoate, which is juvenile hormone III lacking the epoxide group.[7] Methyl farnesoate is believed to play a role similar to that of JH in crustaceans.
Being a sesquiterpenoid, JH chemical structure differs significantly from the structure of other animal hormones. Some JH analogs have been found in conifers.[8]
Control of development
The primary control of juvenile hormone is by 1) the stimulation of the corpora allata by allatotropins[9] short peptides which bind to G-protein coupled receptors,[10] which signal the glands to produce JH, and 2), the inhibition of JH production by allatostatins. These fall into three classes: allatostatin A,[11] allatostatin B,[12] and allatostatin C[13] (for a review of these control mechanisms see: Stay and Woodhead 1993).[14] Secondary control of the JH titre found in the haemolymph of the developing insect is metabolic inactivation of JH by JH-specific esterase and juvenile hormone epoxide hydrolase.[15] During ecdysis the form of the old cuticle laid down before the next moult is controlled by the JH level in the insect. JH maintains a juvenile state. The level gradually decreases during the development of the insect, allowing it to proceed to successive instars with each molt.
This has been demonstrated in various studies, most prominently that by V. B. Wigglesworth in 1960s. In this study, two adult Rhodnius had their blood systems linked, ensuring that the JH titre in both would be equal. One was a third instar Rhodnius, the other was a fourth instar. When the corpora allata of the third instar insect were removed, the level of JH was equal in both insects to that in the fourth instar animal, and hence both proceeded to the fifth instar at the next moult. When the fourth instar Rhodnius had its corpora allata removed, both contained a third instar level of JH and hence one proceeded to instar four, and the other remained at this instar.
Generally, the removal of the corpora allata from juveniles will result in a diminutive adult at the next moult. Implantation of corpora allata into last larval instars will boost JH levels and hence produce a supernumary (extra) juvenile instar etc.
In honey bees
There is a complex interaction between JH, the hormone ecdysone and vitellogenin. In the development stage, as long as there is enough JH, the ecdysone promotes larva-to-larva molts. With lower amounts of JH, ecdysone promotes pupation. Complete absence of JH results in formation of the adult.[16] In adult honey bees, JH and Vitellogenin titers in general show an inverse pattern.[17][18][19][20]
JH titers in worker honey bees progressively increase through the first 15 or so days of the worker's life before the onset of foraging.[21] During the first 15 days, workers perform tasks inside the hive, such as nursing larvae, constructing comb, and cleaning cells. JH titers peak around day 15; workers this age guard, remove dead bees from the colony, and fan at the colony entrance to cool the nest. Aggressiveness of guard bees is correlated with their blood JH levels. Even though guards have high JH levels, their ovaries are relatively undeveloped.[22][23] Although, JH does not activate foraging. Rather it is involved in controlling the pace at which bees develop into foragers.[24]
Vitellogenin titers are high at the beginning of adult life and slowly decrease.
JH has been known to be involved in the queen-worker caste differentiation during the larval stage.[25] The unique negative relationship between JH and Vitellogenin may be important to the understanding of queen longevity.[26]
In Lepidoptera
JH in many butterfly and moth species are necessary for the production and release of the sex pheromone by females. Experiments conducted in Mythimna unipuncta (true armyworm moth) and Agrotis ipsilon (black cutworm moth) have shown that removing the corpus allata, which secretes JH, stops all release of sex pheromone. Furthermore, JH is important for ovarian development.[27][28] In the black cutworm, it was shown that JH is also necessary in males for pheromone responsiveness.[29] JH has also been shown to be transferred from the male to the female Heliothis virescens during copulation.[30]
Forms
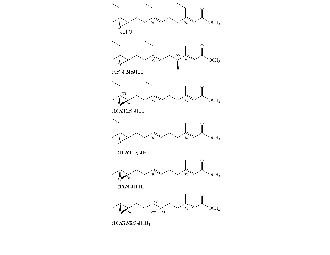
- Methyl farnesoate
- CAS methyl (2E,6E)-3,7,11-trimethyl-2,6,10-dodecatrienoate
- Formula: C16H26O2
- Juvenile hormone 0 (found in Lepidoptera)
- CAS methyl (2E,6E)-10R,11S-(oxiranyl)-3,7-diethyl-11-methyl-2,6-tridecadienoate
- Formula: C19H32O3
- Juvenile hormone I (found in Lepidoptera)
- CAS methyl (2E,6E)-10R,11S-(oxiranyl)-7-ethyl-3,11-dimethyl-2,6-tridecadienoate
- Formula: C18H30O3
- Juvenile hormone II (found in Lepidoptera)
- Juvenile hormone JHB3 (found in diptera)
- CAS methyl (2E,6E)-6S,7S,10R-(dioxiranyl)-3,7,11-trimethyl-2-dodecaenoate
- Formula: C16H26O4
Use as an insecticide
Synthetic analogues of the juvenile hormone are used as an insecticide, preventing the larvae from developing into adult insects. JH itself is expensive to synthesize and is unstable in light. At high levels of JH, larvae can still molt, but the result will only be a bigger larva, not an adult. Thus the insect's reproductive cycle is broken. One JH analogue, methoprene, is approved by the WHO for use in drinking water cisterns to control mosquito larvae due to its exceptionally low toxicity (LD50 >35,000 mg/kg in the rat).
Regulation
Juvenile hormone is produced in the corpora allata of insects. JH will disperse throughout the haemolymph and act on responsive tissues. JH is principally degraded by the enzymes Juvenile-hormone esterase (JHE) or juvenile hormone epoxide hydrolase (JHEH). JHE and JHEH both lead to suppression of JH signaling and response. Tissues responsive to JH may produce one or both of these enzymes.
JH stimulates the accessory glands of adult males, promoting gland growth and the production of accessory gland secretion. Yolk production (vitellogenesis) in female ovaries is also stimulated by JH action. JH may also regulate reproductive behaviour in both sexes.
Insect growth regulators
Insect growth regulators (IGRs) such as juvenile and moulting hormones or their analogs (juvenoids and ecdysoids) when used judiciously, have been found to be useful in insect culture such as sericulture industry. In addition, ecdysoids also show a variety of other uses such as insecticidal, as biochemical tool in gene expression studies, as wound healing and anabolic agents (body building agents with enhancing protein synthesis), as nutraceuticals and cosmetics (hair growth) IGRs occur in insects in very small amounts and are not practical source for these phytochemicals. However, with the discovery of their occurrence in significant quantities in some plants, IGRs and their analogs became easily available in substantial amounts. As a result, many new bioactivities of ecdysoids and juvenoids were discovered. Besides use in insect sericulture, they have found applications in apiculture and aquaculture (prawns). Ecdysoids show remarkable anabolic activities in human and are very much in demand as nutraceuticals (food supplements) including body building agent. Realizing the economic potential of IGRs, bioprospection for these compounds from indigenous plant sources was undertaken. Surveys indicate that a large number plants belonging to different taxa contain IGRs.
Metabolism
Juvenile hormone's activity is destroyed by two enzymes. JH esterase cleaves the methyl ester giving JH acid. JH acid is attached by JH epoxide hydrolase, which converts the epoxide group to a diol. The order of cleavage depends on the order of insect. In the Lepidoptera, the order is as given here; ester cleavage preceding epoxide hydration. Either one terminates the activity of the hormone. JH diol acid, the product of both enzymes, is acted on by JH diol kinase to increase the solubility for excretion.
Biosynthesis
The biosynthesis of JH is similar to that of cholesterol in animals. There are considerable differences between the biosynthesis of the homo-isoprenoid JHs found almost exclusively in Lepidoptera, as opposed to the isoprenoids JH III, JH III bisepoxide, and methyl farnesoate found in other insects.
Cholesterol biosynthesis has been exhaustively studied in animals. All steps occur in the cytosol. The starting material is citrate, which is exported by the mitochondrion when metabolic fuels are high. It is converted into acetyl-CoA, ADP, CO2, and oxaloacetate by ATP-citrate lyase, together with ATP and CoASH as substrates. Three acetyl-CoAs are converted into HMG-CoA by the cytosolic isoforms of thiolase and 3-hydroxy-3-methylglutaryl-CoA synthase. The HMG-CoA is then reduced by NADPH to mevalonate by HMG-CoA reductase, the rate controlling enzyme of cholesterol biosynthesis. This enzyme has 8 helical domains anchoring it in the Golgi membrane of the ER;[31] the catalytic domain is in the cytosol. It is strongly inhibited by the statins, a class of drugs based on a mold metabolite which, at least at one time, were the largest selling class of drugs in the world. Mevalonate is acted of by a series of 3 kinases to give the highly labile 1,2-diphosphomevalonate-3-phosphate, which is acted on by a lyase to give phosphate, CO2, and isopentenyl diphosphate. Isopentenyl diphosphate isomerase converts the latter to the less stable dimethylallyl diphosphate. Farnesyl diphosphate synthase takes one DMAPP and two IPP to give the C15 metabolite farnesyl diphosphate. There are a large number of additional steps to generate cholesterol from IPP, the ubiquitous precursor of all isoprenoids.
It appears that the biosyntheis of JH III is identical to that of cholesterol, from production of IPP to FPP, although there appear to be no studies on export of citrate or other metabolites from the mitochondrion into the cytosol, or formation of acetyl-CoA. The enzymes of this pathway were first studied in Manduca sexta, which produces both homoisoprenoid and isoprenoid (JHIII) JHs.[32]
Very early on propionate was shown to incorporate very highly efficiently into JH II and JH I in cell free extracts of M. sexta] corpora allata.[33][34] Mevalonate and acetate also incorporate into JH I, II, JH III from M. sexta, albeit far less efficiently that propionate.[34] Jennings et al. showed that homomevalonate incorporates into JH II in M. sexta.[35] Baker identified 3-hydroxy-3-ethylglutarate and 3-hydroxy-3-methylglutarate from the same enzyme source incubated with acetyl and propionyl-CoA. [36] Lee et al. showed that the same source of enzymes efficiently make both mevalonate and its 3-ethyl homolog, homomevalonate.[37] Bergot showed that the mevalonate and homomevalonate produced by these enzymes has the same 3S optical isomer configuration as the vertebrate enzymes.[38] Baker showed that isopentenyl diphosphate, and its homolog, 3-ethyl-butenyl diphosphate (homoisopentenyl diphosphate) are metabolized to their corresponding allyic diphosphates, DMAPP and homoDMAPP (3-ethyl-3-methylallyl diphosphate). The latter is required for biosynthesis of JH I, JH II, and 4-methylJH I. 2 units of homoDMAPP are required for JH I and 4methyl JH I biosynthesis, and one for JH II biosynthesis.[39]
All parts of the carbon skeleton comes from IPP. Then an enzyme prenyl transferase/farnesyl diphosphate synthase binds IPP, strips the diphosphate off it to give an allylic carbocation, and adds this to an IPP to give geranyl diphosphate (C10). Then it does the same thing to geranyl diphosphate, giving farnesyl diphosphate (C15). This reaction appears to be the only known enzymatic reaction involving the coupling of two molecules with a carbocation. The free electron pair adds to the double bond of IPP, also isomerizing IPP so that the product is an allylic diphosphate. Thus, this part of the isoprenoid pathway appears nearly identical with that of cholesterol with the exception of the insect specific homoisoprenoid units. NAD+-dependent farnesol dehydrogenase, a corpora allata enzyme involved in juvenile hormone synthesis showed that the same source of enzymes efficiently make both mevalonate and its 3-ethyl homolog, homomevalonate.[40]
Absolute configuration of homomevalonate and 3-hydroxy-3-ethylglutaryl and 3-hydroxy-3-methylglutaryl coenzyme a, produced by cell-free extracts of insect corpora allata. A cautionary note on prediction of absolute stereochemistry based on liquid chromatographic elution order of diastereomeric derivatives.[41] showed that the mevalonate and homomevalonate produced by these enzymes has the same 3S optical isomer configuration as the vertebrate enzymes[39] showed that isopentenyl diphosphate, and its homolog, 3-ethyl-butenyl diphosphate (homoisopentenyl diphosphate) are metabolized to their corresponding allyic diphosphates, DMAPP and homoDMAPP (3-ethyl-3-methylallyl diphosphate). The latter is required for biosynthesis of JH I, JH II, and 4-methylJH I. 2 units of homoDMAPP are required for JH I and 4methyl JH I biosynthesis, and one for JH II biosynthesis.
However, at this point these pathways diverge. While the vast majority of farnesyl diphosphate is converted into ultimately cholesterol in animals, in insects it is apparently acting on by a diphosphatase to give farnesol, which in turn is acted on by an NAD+ dependent enzyme, farnesol/farnesal dehydrogenase in M. sexta[40] to give farnesoic acid. Subsequent work has shown that the enzyme is highly specific for trans allylic alcohols with at least three isoprene units,[42] and to also be present in mosquitoes.[43]
The next steps of JH biosynthesis differ between orders. In Lepidoptera and mosquitoes farnesoic acid or its homologs is epoxidized by a P450 dependent farnesoic acid methyl epoxidase, then it is methylated by a JH acid methyl transferase[44] In most orders, farnesoic acid is methylated by farensoic acid methyl transferase, and then is epoxidized by a P450 dependent methyl transferas.[44]
A recent publication by Nouzova et al. (2015) shows that allatostatin C, the peptide which inhibits JH production by the corpora allata, blocks the transport of citrate out of the mitochondrion in Aedes aegypti. This is a very logical control mechanism for JH biosynthesis.[45]
References
- Riddiford, L. M. (1994). "Cellular and molecular actions of juvenile hormone I. General considerations and premetamorphic actions". Advances in Insect Physiology. 24: 213–274. doi:10.1016/S0065-2806(08)60084-3. ISBN 9780120242245.
- Watt, G. R.; Davey, K. G. (1996). "Cellular and Molecular Actions of Juvenile Hormone. II. Roles of Juvenile Hormone in Adult Insects". Advances in Insect Physiology. 26: 1–155. doi:10.1016/S0065-2806(08)60030-2. ISBN 9780120242269.
- Nijhout, H. F. (1994). Insect Hormones. Princeton: Princeton University Press.
- Röller, H.; Dahm, K.H.; Sweeley, C.C.; Trost, B.M. (1967). "The Structure of the Juvenile Hormone". Angewandte Chemie International Edition. 6 (2): 179–180. doi:10.1002/anie.196701792.
- Judy, K.J.; Schooley, D.A.; Dunham, L.L.; Hall, M.S.; Bergot, B.J.; Siddall, J.B. (1973). "Isolation, Structure, and Absolute Configuration of a New Natural Insect Juvenile Hormone from Manduca sexta". Proceedings of the National Academy of Sciences of the United States of America. 70 (5): 1509–1513. Bibcode:1973PNAS...70.1509J. doi:10.1073/pnas.70.5.1509. PMC 433531. PMID 16592086.
- Richard, D.S.; Applebaum, S.W.; Sliter, T.J.; Baker, F.C.; Schooley, D.A.; Reuter, C.C.; Henrich, V.C.; Gilbert, L.I. (1989). "Juvenile Hormone Bisepoxide Biosynthesis in vitro by the Ring Gland of Drosophila melanogaster: A putative juvenile Hormone in the higher Diptera". Proceedings of the National Academy of Sciences of the United States of America. 86 (4): 1421–1425. Bibcode:1989PNAS...86.1421R. doi:10.1073/pnas.86.4.1421. PMC 286704. PMID 2493154.
- Laufer, H.; Borst, D.; Baker, F.C.; Carasco, C.; Sinkus, M.; Reuter, C.C.; Tsai, L.W.; Schooley, D.A. (1987). "Identification of a juvenile hormone-like compound in a crustacean". Science. 235 (4785): 202–205. Bibcode:1987Sci...235..202L. doi:10.1126/science.235.4785.202. PMID 17778635.
- Rogers, I. H.; J. F. Manville; T. Sahota (April 1974). "Juvenile Hormone Analogs in Conifers. II. Isolation, Identification, and Biological Activity of cis-4-[1′(R)-5′-Dimethyl-3′-oxohexyl]-cyclohexane-1-carboxylic Acid and (+)-4(R)-[1′(R)-5′-Dimethyl-3′-oxohexyl]-1-cyclohexene-1-carboxylic Acid from Douglas-fir Wood". Canadian Journal of Chemistry. 52 (7): 1192–1199. doi:10.1139/v74-187.
- Schooley, D.A., 1991. Identification of an allatostatin from the tobacco hornworm Manduca sexta. Proceedings of the National Academy of Sciences of the United States of America 88, 9458-9462
- Cusson, M.; Prestwich, G.D.; Stay, B.; Tobe, S.S. (1991). "Photoaffinity labeling of allatostatin receptor proteins in the corpora allata of the cockroach, Diploptera punctata". Biochemical and Biophysical Research Communications. 181 (2): 736–742. doi:10.1016/0006-291x(91)91252-8. PMID 1661589.
- Woodhead, A.P.; Stay, B.; Seidel, S.L.; Khan, M.A.; Tobe, S.S. (1989). "Primary structure of four allatostatins: Neuropeptide inhibitors of juvenile hormone biosynthesis". Proceedings of the National Academy of Sciences of the United States of America. 86 (15): 5997–6001. Bibcode:1989PNAS...86.5997W. doi:10.1073/pnas.86.15.5997. PMC 297759. PMID 2762309.
- Reichwald, K.; Unnithan, G.C.; Davis, N.T.; Agricola, H.; Feyereisen, R. (1994). "Expression of the allatostatin gene in endocrine cells of the cockroach midgut". Proceedings of the National Academy of Sciences of the United States of America. 91 (25): 11894–11898. Bibcode:1994PNAS...9111894R. doi:10.1073/pnas.91.25.11894. PMC 45342. PMID 7991553.
- Kramer, S.J.; Toschi, A.; Miller, C.A.; Kataoka, H.; Quistad, G.B.; Li, J.P.; Carney, R.L.; Schooley, D.A. (1991). "Identification of an allatostatin from the tobacco hornworm Manduca sexta". Proceedings of the National Academy of Sciences of the United States of America. 88 (21): 9458–9462. Bibcode:1991PNAS...88.9458K. doi:10.1073/pnas.88.21.9458. PMC 52737. PMID 1946359.
- Stay, B.; Woodhead, A.P. (1993). "Neuropeptide regulators of insect corpora allata". American Zoologist. 33 (3): 357–364. doi:10.1093/icb/33.3.357.
- White, A.F. (1972). "Metabolism of the Juvenile Hormone Analogue Methyl Farnesoate 10,11-Epoxide in Two Insect Species". Life Sciences. 11 (4): 201–210. doi:10.1016/0024-3205(72)90110-5.
- Kimball, John W. (2002) Insect hormones Archived 3 January 2010 at the Wayback Machine
- Hartfelder K, Engels W (1998) Curr Top Dev Biol 40:45–77
- Bloch G, Wheeler DE, Robinson GE (2002) in Hormones, Brain, and Behavior, ed Pfaff D (Academic, New York) Vol 3, 195–236
- Fluri P, Sabatini AG, Vecchi MA, Wille H (1981) J Apic Res 20:221–225
- Fahrbach SE, Giray T, Robinson GE (1995) Neurobiol Learn Mem 63:181–191
- Elekonich, M. M., Schulz, D. J., Bloch, G. and Robinson, G. E. (2001). Juvenile hormone levels in honey bee (Apis mellifera L.) foragers: foraging experience and diurnal variation. Journal for Insect Physiology. 47,1119 -1125
- Pearce A.N., Huang Z.Y., Breed M.D. (2001) Juvenile hormone and aggression in honey bees, Journal for Insect Physiology. 47, 1243– 1247.
- Breed, Michael D. (2002) Juvenile Hormone
- Sullivan, J. P., Jassim, O., Fahrbach, S. E. and Robinson, G. E. (2000). Juvenile hormone paces behavioral development in the adult worker honey bee. Hormones and Behavior. 37, 1-14
- Rachinsky A., Hartfelder K. (1990) Corpora allata activity, a primer regulating element for caste juvenile hormone titer in honey bee larvae (Apis mellifera carnica), Journal of Insect Physiology. 36, 189–194
- Miguel Corona, Rodrigo A. Velarde, Silvia Remolina, Adrienne Moran-Lauter, Ying Wang, Kimberly A. Hughes, and Gene E. Robinson Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity Proceedings of the National Academy of Sciences of the United States of America, April 2007; 104: 7128 - 7133
- Cusson, M., and J. N. Mcneil. "Involvement of Juvenile Hormone in the Regulation of Pheromone Release Activities in a Moth." Science, vol. 243, no. 4888, 1989, pp. 210–212., doi:10.1126/science.243.4888.210.
- Picimbon, Jean-Francois. "Juvenile Hormone Stimulates Pheromonotropic Brain Factor Release in the Female Black Cutworm, Agrotis Ipsilon". Journal of Insect Physiology, vol. 41, no. 5, pp. 377–382.
- Gadenne, C., et al. "Hormonal Control of Pheromone Responsiveness in the Male Black Cutworm Agrotis ipsilon." Experientia, vol. 49, no. 8, 1993, pp. 721–724., doi:10.1007/bf01923960.
- Park, Yong Ihl (1998). "Mating in Heliothis virescens: Transfer of Juvenile Hormone During Copulation by Male to Female and Stimulation of Biosynthesis of Endogenous Juvenile Hormone". Archives of Insect Biochemistry and Physiology. 38 (2): 100–107. doi:10.1002/(SICI)1520-6327(1998)38:2<100::AID-ARCH6>3.0.CO;2-X. PMID 9627408.
- Burg, J.S., Espenshade, P.J., 2011. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 50, 403-410
- Judy, K.J.; Schooley, D.A.; Dunham, L.L.; Hall, M.S.; Bergot, B.J.; Siddall, J.B. (1973). "Isolation, Structure, and Absolute Configuration of a New Natural Insect Juvenile Hormone from Manduca sexta". Proceedings of the National Academy of Sciences of the United States of America. 70 (5): 1509–1513. Bibcode:1973PNAS...70.1509J. doi:10.1073/pnas.70.5.1509. PMC 433531. PMID 16592086.
- Peter, M.G.; Dahm, K.H. (1975). "Biosynthesis of Juvenile Hormone in the Cecropia Moth. Labelling Pattern from [1-14+C]-Propionate through Degradation to Single Carbon Atom Derivatives". Helvetica Chimica Acta. 58 (4): 1037–1048. doi:10.1002/hlca.19750580407. PMID 1158736.
- Schooley, D.A.; Judy, K.J.; Bergot, B.J.; Hall, M.S.; Siddall, J.B. (1973). "Biosynthesis of the Juvenile Hormones of Manduca sexta: Labeling Pattern from Mevalonate, Propionate, and Acetate". Proceedings of the National Academy of Sciences of the United States of America. 70 (10): 2921–2925. Bibcode:1973PNAS...70.2921S. doi:10.1073/pnas.70.10.2921. PMC 427139. PMID 16592112.
- Jennings, Richard C.; Judy, Kenneth J.; Schooley, David A. (1975). "Biosynthesis of the homosesquiterpenoid juvenile hormone JH II [methyl (2E,6E,10Z)-10,11 -epoxy-3,7,11-trimethyltridecadienoate] from [5-3H]homomevalonate in Manduca sexta". Journal of the Chemical Society, Chemical Communications (1): 21. doi:10.1039/c39750000021. ISSN 0022-4936.
- Baker, Fred. C.; Schooley, David A. (1978). "Juvenile hormone biosynthesis: identification of 3-hydroxy-3-ethylglutarate and 3-hydroxy-3-methylglutarate in cell-free extracts from Manduca sexta incubated with propionyl- and acetyl-CoA". Journal of the Chemical Society, Chemical Communications (7): 292. doi:10.1039/c39780000292. ISSN 0022-4936.
- Lee, Eun; Schooley, David A.; Hall, M. Sharon; Judy, Kenneth J. (1978). "Juvenile hormone biosynthesis: homomevalonate and mevalonate synthesis by insect corpus allatum enzymes". Journal of the Chemical Society, Chemical Communications (7): 290. doi:10.1039/c39780000290. ISSN 0022-4936.
- Bergot, B. John; Baker, Frederick Charles; Lee, Eun; Schooley, David A. (November 1979). "Absolute configuration of homomevalonate and 3-hydroxy-3-ethylglutaryl- and 3-hydroxy-3-methylglutaryl CoA, produced by cell-free extracts of insect corpora allata; cautionary note on prediction of absolute stereochemistry based on liquid chromatographic elution order of diastereomeric derivatives". Journal of the American Chemical Society. 101 (24): 7432–7434. doi:10.1021/ja00518a064. ISSN 0002-7863.
- Baker, F.C., Lee, E., Bergot, B.J., Schooley, D.A., 1981. Isomerization of isopentenyl pyrophosphate and homoisopentenyl pyrophosphate by Manduca sexta corpora cardiaca - corpora allata homogenates., in: Pratt, G.E., Brooks, G.T. (Eds.), Juvenile Hormone Biochemistry. Elsevier, Amsterdam, pp. 67-80
- Baker, F.C.; Mauchamp, B.; Tsai, L.W.; Schooley, D.A. (1983). "Farnesol and farnesal dehydrogenase(s) in corpora allata of the Tobacco Hornworm moth, Manduca sexta". Journal of Lipid Research. 24 (12): 1586–1594. PMID 6366103.
- Journal of the American Chemical Society 101, 7432-7434
- Sen, S.E.; Garvin, G.M. (1995). "Substrate requirements for lepidopteran farnesol dehydrogenase". Journal of Agricultural and Food Chemistry. 43 (3): 820–825. doi:10.1021/jf00051a049.
- Mayoral, J.G.; Nouzova, M.; Navare, A.; Noriega, F.G. (2009). "NAD+-dependent farnesol dehydrogenase, a corpora allata enzyme involved in juvenile hormone synthesis". Proceedings of the National Academy of Sciences of the United States of America. 106 (50): 21091–21096. Bibcode:2009PNAS..10621091M. doi:10.1073/pnas.0909938106. PMC 2795498. PMID 19940247.
- Defelipe, L.A.; Dolghih, E.; Roitberg, A.E.; Nouzova, M.; Mayoral, J.G.; Noriega, F.G.; Turjanski, A.G. (2011). "Juvenile hormone synthesis: "esterify then epoxidize" or "epoxidize then esterify"? Insights from the structural characterization of juvenile hormone acid methyltransferase". Insect Biochemistry and Molecular Biology. 41 (4): 228–235. doi:10.1016/j.ibmb.2010.12.008. PMC 3057355. PMID 21195763.
- Nouzova, Marcela; Rivera-Perez, Crisalejandra; Noriega, Fernando G. (February 2015). "Allatostatin-C reversibly blocks the transport of citrate out of the mitochondria and inhibits juvenile hormone synthesis in mosquitoes". Insect Biochemistry and Molecular Biology. 57: 20–26. doi:10.1016/j.ibmb.2014.12.003. PMC 4293212. PMID 25500428.
Further reading
- Wigglesworth, VB. (1964) The hormonal regulation of growth and reproduction in insects. Adv. Insect Physiol. 2: 247-336
- Wigglesworth, VB. (1939) The Principles of Insect Physiology. Cambridge University Press. Cambridge.
- Berger and Dubrovsky (2005) Juvenile Hormone Molecular Actions and Interactions During Development of Drosophila. Vitamins and Hormones. 73:172-215