Toxoplasma gondii
Toxoplasma gondii (/ˈtɒksoʊplæzmə ˈɡɒndiaɪ/) is an obligate intracellular parasitic one-celled eukaryote (specifically an apicomplexan) that causes the infectious disease toxoplasmosis.[3] Found worldwide, T. gondii is capable of infecting virtually all warm-blooded animals,[4]:1 but felids, such as domestic cats, are the only known definitive hosts in which the parasite may undergo sexual reproduction.[5][6]
| Toxoplasma gondii | |
|---|---|
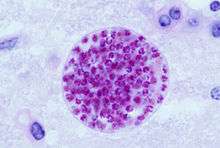
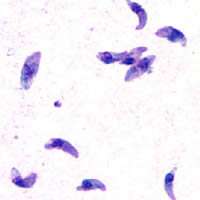 | |
| Giemsa stained T. gondii tachyzoites, 1000× magnification | |
| Scientific classification | |
| Kingdom: | Chromista |
| Infrakingdom: | Alveolata |
| Phylum: | Apicomplexa |
| Class: | Conoidasida |
| Order: | Eucoccidiorida |
| Family: | Sarcocystidae |
| Subfamily: | Toxoplasmatinae |
| Genus: | Toxoplasma Nicolle & Manceaux, 1909[1] |
| Species: | T. gondii |
| Binomial name | |
| Toxoplasma gondii (Nicolle & Manceaux, 1908)[2] | |
In humans, T. gondii is one of the most common parasites in developed countries;[7][8] serological studies estimate that 30–50% of the global population has been exposed to and may be chronically infected with T. gondii, although infection rates differ significantly from country to country.[9][10] For example, estimates have shown the highest prevalence of persons infected to be in France, at 84%, as at 2000.[11] Although mild, flu-like symptoms occasionally occur during the first few weeks following exposure, infection with T. gondii produces no readily observable symptoms in healthy human adults.[9][12][4]:77 This asymptomatic state of infection is referred to as a latent infection and has recently been associated with numerous subtle adverse or pathological behavioral alterations in humans,[9][13] though it has been shown recently that the association between behavioural changes and infection with T. gondii is weak.[14] In infants, HIV/AIDS patients, and others with weakened immunity, infection may cause a serious and occasionally fatal illness, toxoplasmosis.[12][4]:77
T. gondii has been shown to alter the behavior of infected rodents in ways that increase the rodents' chances of being preyed upon by felids.[11][15][16] Support for this "manipulation hypothesis" stems from studies showing T. gondii-infected rats have a decreased aversion to cat urine.[11] Because cats are the only hosts within which T. gondii can sexually reproduce to complete and begin its lifecycle, such behavioral manipulations are thought to be evolutionary adaptations that increase the parasite's reproductive success.[11] The rats would not shy away from areas where cats live and would also be less able to escape should a cat try to prey on them. The primary mechanisms of T. gondii–induced behavioral changes in rodents is now known to occur through epigenetic remodeling in neurons which govern the associated behaviors;[17][18] for example, it modifies epigenetic methylation to cause hypomethylation of arginine vasopressin-related genes in the medial amygdala to greatly decrease predator aversion.[17][18] Widespread histone-lysine acetylation in cortical astrocytes appears to be another epigenetic mechanism employed by T. gondii.[19][20] Differences in aversion to cat urine are observed between non-infected and infected humans and sex differences within these groups were apparent, too.[21]
A number of studies have suggested that subtle behavioral or personality changes may occur in infected humans,[22] and infection with the parasite has recently been associated with a number of neurological disorders, particularly schizophrenia[16] and bipolar disorder.[23][24] A 2015 study also found cognitive deficits in adults to be associated with joint infection by both T. gondii and Helicobacter pylori in a regression model with controls for race-ethnicity and educational attainment.[25] Although a causal relationship between latent toxoplasmosis with these neurological phenomena has not yet been established,[9][16] preliminary evidence suggests that T. gondii infection may induce some of the same alterations in the human brain as those observed in mice.[26][27]
Structure
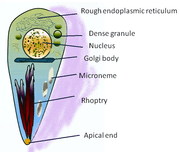
T. gondii contains, amongst other organelles, rhoptry and microneme.
Lifecycle
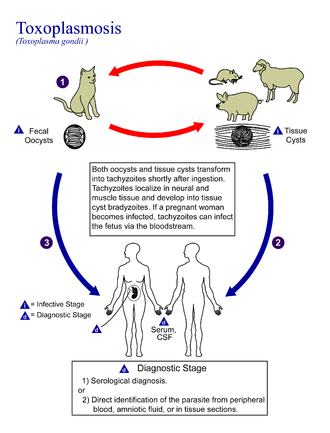

The lifecycle of T. gondii may be broadly summarized into two components: a sexual component that occurs only within cats (felids, wild or domestic), and an asexual component that can occur within virtually all warm-blooded animals, including humans, cats, and birds.[28]:2 Because T. gondii can sexually reproduce only within cats, cats are therefore the definitive host of T. gondii. All other hosts – in which only asexual reproduction can occur – are intermediate hosts.
Sexual reproduction in the feline definitive host
When a feline is infected with T. gondii (e.g. by consuming an infected mouse carrying the parasite's tissue cysts), the parasite survives passage through the stomach, eventually infecting epithelial cells of the cat's small intestine.[28]:39 Inside these intestinal cells, the parasites undergo sexual development and reproduction, producing millions of thick-walled, zygote-containing cysts known as oocysts. Felines are the only definitive host because they lack expression of the enzyme delta-6-desaturase (D6D) in their intestine. This enzyme converts linoleic acid; the absence of expression allows systemic linoleic acid accumulation. Recent findings showed that this excess of linoleic acid is essential for T. gondii sexual reproduction.[6]
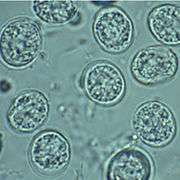
Feline shedding of oocysts
Infected epithelial cells eventually rupture and release oocysts into the intestinal lumen, whereupon they are shed in the cat's feces.[4]:22 Oocysts can then spread to soil, water, food, or anything potentially contaminated with the feces. Highly resilient, oocysts can survive and remain infective for many months in cold and dry climates.[29]
Ingestion of oocysts by humans or other warm-blooded animals is one of the common routes of infection.[30] Humans can be exposed to oocysts by, for example, consuming unwashed vegetables or contaminated water, or by handling the feces (litter) of an infected cat.[28]:2[31] Although cats can also be infected by ingesting oocysts, they are much less sensitive to oocyst infection than are intermediate hosts.[32][4]:107
Initial infection of the intermediate host
T. gondii is considered to have three stages of infection; the tachyzoite stage of rapid division, the bradyzoite stage of slow division within tissue cysts, and the oocyst environmental stage.[33] Tachyzoites are also known as "tachyzoic merozoites" and bradyzoites as "bradyzoic merozoites".[34] When an oocyst or tissue cyst is ingested by a human or other warm-blooded animal, the resilient cyst wall is dissolved by proteolytic enzymes in the stomach and small intestine, freeing sporozoites from within the oocyst.[30][33] The parasites first invade cells in and surrounding the intestinal epithelium, and inside these cells, the parasites differentiate into tachyzoites, the motile and quickly multiplying cellular stage of T. gondii.[28]:39 Tissue cysts in tissues such as brain and muscle tissue, form about 7–10 days after initial infection.[33]
Asexual reproduction in the intermediate host
Inside host cells, the tachyzoites replicate inside specialized vacuoles (called the parasitophorous vacuoles) created during parasitic entry into the cell.[28]:23–39 Tachyzoites multiply inside this vacuole until the host cell dies and ruptures, releasing and spreading the tachyzoites via the bloodstream to all organs and tissues of the body, including the brain.[28]:39–40
Growth in tissue culture
The parasite can be easily grown in monolayers of mammalian cells maintained in vitro in tissue culture. It readily invades and multiplies in a wide variety of fibroblast and monocyte cell lines. In infected cultures, the parasite rapidly multiplies and thousands of tachyzoites break out of infected cells and enter adjacent cells, destroying the monolayer in due course. New monolayers can then be infected using a drop of this infected culture fluid and the parasite indefinitely maintained without the need of animals.
Formation of tissue cysts
Following the initial period of infection characterized by tachyzoite proliferation throughout the body, pressure from the host's immune system causes T. gondii tachyzoites to convert into bradyzoites, the semidormant, slowly dividing cellular stage of the parasite.[35] Inside host cells, clusters of these bradyzoites are known as tissue cysts. The cyst wall is formed by the parasitophorous vacuole membrane.[28]:343 Although bradyzoite-containing tissue cysts can form in virtually any organ, tissue cysts predominantly form and persist in the brain, the eyes, and striated muscle (including the heart).[28]:343 However, specific tissue tropisms can vary between intermediate host species; in pigs, the majority of tissue cysts are found in muscle tissue, whereas in mice, the majority of cysts are found in the brain.[28]:41
Cysts usually range in size between five and 50 µm in diameter,[36] (with 50 µm being about two-thirds the width of the average human hair).[37]
Consumption of tissue cysts in meat is one of the primary means of T. gondii infection, both for humans and for meat-eating, warm-blooded animals.[28]:3 Humans consume tissue cysts when eating raw or undercooked meat (particularly pork and lamb).[38] Tissue cyst consumption is also the primary means by which cats are infected.[4]:46
An exhibit at the San Diego Natural History Museum states urban runoff with cat feces transports Toxoplasma gondii into the ocean, which can kill sea otters.[39]
Chronic infection
Tissue cysts can be maintained in host tissue for the lifetime of the animal.[28]:580 However, the perpetual presence of cysts appears to be due to a periodic process of cyst rupturing and re-encysting, rather than a perpetual lifespan of individual cysts or bradyzoites.[28]:580 At any given time in a chronically infected host, a very small percentage of cysts are rupturing,[28]:45 although the exact cause of this tissue cysts rupture is, as of 2010, not yet known.[4]:47
Theoretically, T. gondii can be passed between intermediate hosts indefinitely via a cycle of consumption of tissue cysts in meat. However, the parasite's lifecycle begins and completes only when the parasite is passed to a feline host, the only host within which the parasite can again undergo sexual development and reproduction.[30]
Population structure in the wild
Khan et al.[40] reviewed evidence that despite the occurrence of a sexual phase in its life cycle, T. gondii has an unusual population structure dominated by three clonal lineages (Types I, II and III) that occur in North America and Europe. They estimated that a common ancestor founded these about 10,000 years ago. In a further and larger study (with 196 isolates from diverse sources including T. gondii found in the bald eagle, gray wolves, Arctic foxes and sea otters), Dubey et al.[41] also found that T. gondii strains infecting North American wildlife have limited genetic diversity with the occurrence of only a few major clonal types. They found that 85% of strains in North America were of one of three widespread genotypes (Types II, III and Type 12). Thus T. gondii has retained the capability for sex in North America over many generations, producing largely clonal populations, and matings have generated little genetic diversity.
Cellular stages
During different periods of its life cycle, individual parasites convert into various cellular stages, with each stage characterized by a distinct cellular morphology, biochemistry, and behavior. These stages include the tachyzoites, merozoites, bradyzoites (found in tissue cysts), and sporozoites (found in oocysts).
Tachyzoites
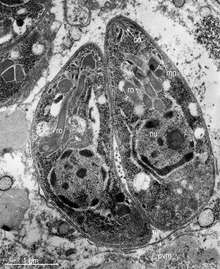
Motile, and quickly multiplying, tachyzoites are responsible for expanding the population of the parasite in the host.[42][28]:19 When a host consumes a tissue cyst (containing bradyzoites) or an oocyst (containing sporozoites), the bradyzoites or sporozoites stage-convert into tachyzoites upon infecting the intestinal epithelium of the host.[28]:359 During the initial acute period of infection, tachyzoites spread throughout the body via the blood stream.[28]:39–40 During the later, latent (chronic) stages of infection, tachyzoites stage-convert to bradyzoites to form tissue cysts.
Merozoites
Like tachyzoites, merozoites divide quickly, and are responsible for expanding the population of the parasite inside the cat's intestine before sexual reproduction.[28]:19 When a feline definitive host consumes a tissue cyst (containing bradyzoites), bradyzoites convert into merozoites inside intestinal epithelial cells. Following a brief period of rapid population growth in the intestinal epithelium, merozoites convert into the noninfectious sexual stages of the parasite to undergo sexual reproduction, eventually resulting in zygote-containing oocysts.[28]:306
Bradyzoites
Bradyzoites are the slowly dividing stage of the parasite that make up tissue cysts. When an uninfected host consumes a tissue cyst, bradyzoites released from the cyst infect intestinal epithelial cells before converting to the proliferative tachyzoite stage.[28]:359 Following the initial period of proliferation throughout the host body, tachyzoites then convert back to bradyzoites, which reproduce inside host cells to form tissue cysts in the new host.
Sporozoites
Sporozoites are the stage of the parasite residing within oocysts. When a human or other warm-blooded host consumes an oocyst, sporozoites are released from it, infecting epithelial cells before converting to the proliferative tachyzoite stage.[28]:359
Immune response
Initially, a T. gondii infection stimulates production of IL-2 and IFN-γ by the innate immune system.[35] Continuous IFN-γ production is necessary for control of both acute and chronic T. gondii infection.[35] These two cytokines elicit a CD4+ and CD8+ T-cell mediated immune response.[35] Thus, T-cells play a central role in immunity against Toxoplasma infection. T-cells recognize Toxoplasma antigens that are presented to them by the body's own Major Histocompatibility Complex (MHC) molecules. The specific genetic sequence of a given MHC molecule differs dramatically between individuals, which is why these molecules are involved in transplant rejection. Individuals carrying certain genetic sequences of MHC molecules are much more likely to be infected with Toxoplasma. One study of >1600 individuals found that Toxoplasma infection was especially common among people who expressed certain MHC alleles (HLA-B*08:01, HLA-C*04:01, HLA-DRB 03:01, HLA-DQA*05:01 and HLA-DQB*02:01).[43]
IL-12 is produced during T. gondii infection to activate natural killer (NK) cells.[35] Tryptophan is an essential amino acid for T. gondii, which it scavenges from host cells. IFN-γ induces the activation of indole-amine-2,3-dioxygenase (IDO) and tryptophan-2,3-dioxygenase (TDO), two enzymes that are responsible for the degradation of tryptophan.[44] Immune pressure eventually leads the parasite to form cysts that normally are deposited in the muscles and in the brain of the hosts.[35]
Immune response and behaviour alterations
The IFN-γ-mediated activation of IDO and TDO is an evolutionary mechanism that serves to starve the parasite, but it can result in depletion of tryptophan in the brain of the host. IDO and TDO degrade tryptophan to N-formylkynurenine and administration of L-kynurenine is capable of inducing depressive-like behaviour in mice.[44] T. gondii infection has been demonstrated to increase the levels of kynurenic acid (KYNA) in the brains of infected mice and KYNA has also been demonstrated to be increased in the brain of schizophrenic persons.[44] Low levels of tryptophan and serotonin in the brain were already associated to depression.[45]
Risk factors for human infection
The following have been identified as being risk factors for T. gondii infection in humans and warm-blooded animals:
- by consuming raw or undercooked meat containing T. gondii tissue cysts.[31][46][47][48][49] The most common threat to citizens in the United States is from eating raw or undercooked pork.[50]
- by ingesting water, soil, vegetables, or anything contaminated with oocysts shed in the feces of an infected animal.[46] Cat fecal matter is particularly dangerous: Just one cyst consumed by a cat can result in thousands of oocysts. This is why physicians recommend pregnant or ill persons do not clean the cat's litter box at home.[50] These oocysts are resilient to harsh environmental conditions and can survive over a year in contaminated soil.[33][51]
- from a blood transfusion or organ transplant[52]
- from transplacental transmission from mother to fetus, particularly when T. gondii is contracted during pregnancy[46]
- from drinking unpasteurized goat milk[47]
- from raw and treated sewage and bivalve shellfish contaminated by treated sewage[53][54][55][56]
Cleaning cat litter boxes is a potential route of infection;[31] however, numerous studies have shown living in a household with a cat is not a significant risk factor for T. gondii infection,[31][48][57] though living with several kittens has some significance.[58]
In warm-blooded animals, such as brown rats, sheep, and dogs, T. gondii has also been shown to be sexually transmitted.[59][60][61] Although T. gondii can infect, be transmitted by, and asexually reproduce within humans and virtually all other warm-blooded animals, the parasite can sexually reproduce only within the intestines of members of the cat family (felids).[30] Felids are therefore the definitive hosts of T. gondii; all other hosts (like human or other mammals) are intermediate hosts.
Preventing infection
The following precautions are recommended to prevent or greatly reduce the chances of becoming infected with T. gondii. This information has been adapted from the websites of United States Centers for Disease Control and Prevention[62] and the Mayo Clinic.[63]
From food
Basic food-handling safety practices can prevent or reduce the chances of becoming infected with T. gondii, such as washing unwashed fruits and vegetables, and avoiding raw or undercooked meat, poultry, and seafood. Other unsafe practices such as drinking unpasteurized milk or untreated water can increase odds of infection.[62] As T. gondii is commonly transmitted through ingesting microscopic cysts in the tissues of infected animals, meat that is not prepared to destroy these presents a risk of infection. Freezing meat for several days at subzero temperatures (0 °F or −18 °C) before cooking may break down all cysts, as they rarely survive these temperatures.[4]:45 During cooking, whole cuts of red meat should be cooked to an internal temperature of at least 145 °F (63 °C). Medium rare meat is generally cooked between 130 and 140 °F (55 and 60 °C),[64] so cooking meat to at least medium is recommended. After cooking, a rest period of 3 min should be allowed before consumption. However, ground meat should be cooked to an internal temperature of at least 160 °F (71 °C) with no rest period. All poultry should be cooked to an internal temperature of at least 165 °F (74 °C). After cooking, a rest period of 3 min should be allowed before consumption.
From environment
Oocysts in cat feces take at least a day to sporulate (to become infectious after they are shed), so disposing of cat litter daily greatly reduces the chance of infectious oocysts developing. As these can spread and survive in the environment for months, humans should wear gloves when gardening or working with soil, and should wash their hands promptly after disposing of cat litter. These precautions apply to outdoor sandboxes/play sand pits, which should be covered when not in use. Cat feces should never be flushed down a toilet.
Pregnant women are at higher risk of transmitting the parasite to their unborn child and immunocompromised people of acquiring a lingering infection. Because of this, they should not change or handle cat litter boxes. Ideally, cats should be kept indoors and fed only food that has low to no risk of carrying oocysts, such as commercial cat food or well-cooked table food.
Vaccination
As of 2016, no approved human vaccine exists against Toxoplasma gondii.[65] Research on human vaccines is ongoing.[66]
For sheep, an approved live vaccine sold as Toxovax (from MSD Animal Health) provides lifetime protection.[67]
Environmental effects
In many parts of the world, where there are high populations of feral cats, there is an increased risk to the native wildlife due to increased infection of Toxoplasma gondii. It has been found that the serum concentrations of T. gondii in the wildlife population were increased where there are high amounts of cat populations. Therefore, creating an incredibly dangerous environment when the organism has not evolved in cohabitation with felines and their contributing parasites [68]
Impact on marine species
Minks and otters
Toxoplasmosis is one of the contributing factors towards mortality in southern sea otters, especially in areas where there is large urban run-off.[69] In their natural habitats, sea otters control sea urchin populations and, thus indirectly, control sea kelp forests. By enabling the growth of sea kelp, other marine populations are protected as well as CO2 emissions are reduced due to the kelps ability to absorb atmospheric carbon.[70] An examination on 105 beachcast otters revealed that 38.1% had parasitic infections, and 28% of said infections had resulted in protozoal meningoencephalitis deaths.[69] Toxoplasma gondii was found to be the root cause in 16.2% of these deaths, while 6.7% of the deaths were due to a closely related protozoan parasite known as Sarcocystis neurona.[69]
Minks, being semiaquatic, are also susceptible to infection and be antibody positive towards Toxoplasma gondii.[71] Minks can follow a similar diet as otters and feasts on crustaceans, fish, and invertebrates, thus the transmission route follows a similar pattern to otters. Because of the mink's ability to transverse land more frequently, and often seen as an invasive species itself, minks are a bigger threat in transporting T. gondii to other mammalian species, rather than otters who have a more restrictive breadth.[71]
Black-footed penguins
Although under-studied, penguin populations, especially those that share an environment with the human population, are at-risk due to parasite infections, mainly Toxoplasmosis gondii. The main subspecies of penguins found to be infected by T. gondii include wild Magellanic and Galapagos penguins, as well as blue and African penguins in captivity.[72] In one study, 57 (43.2%) of 132 serum samples of Magellanic penguins were found to have T. gondii. The island that the penguin is located, Magdalena Island, is known to have no cat populations, but a very frequent human population, indicating the possibility of transmission.[72]
Histopathology
Examination of black-footed penguins with toxoplasmosis reveals hepatomegaly, splenomegaly, cranial hemorrhage, and necrotic kidneys (Ploeg, et al., 2011). Alveolar and hepatic tissue presents a high number of immune cells like macrophages containing tachyzoites of T. gondii.[73] Histopathological features in other animals affected with toxoplasmosis had tachyzoites in eye structures like the retina which lead to blindness.[73]
Water transmission
The transmission of oocysts has been unknown, even though there have many documented cases of infection in marine species. Researchers have found that the oocytes of T. gondii can survive in seawater for at least 6 months, with the amount of salt concentration not affecting its lifecycle. There have been no studies on the ability of T. gondii oocysts lifecycle within freshwater environments, though infections are still present. One possible hypothesis of transmission is via amoeba species, particularly Acanthamoeba spp, a species that is found in all water environments (fresh-, brackish, and full-strength seawater). Normally, amoebas’ function as a natural filter, phagocytizing nutrients and bacteria found within the water. Some pathogens have used this to their advantage, however, and evolved to be able to avoid being broken down and, thus, survive encased in the amoeba – this includes Holosporaceae, Pseudomonaceae, Burkholderiacceae, among others.[74] Overall, this aids the pathogen in transportation but, also, protection from drugs and sterilizers that would, otherwise, cause death in the pathogen.[75] Studies have shown that T. gondii oocysts can live within amoebas after being engulfed for at least 14 days without significant obliteration of the parasite.[76] The ability of the microorganism to survive in vitro is dependent on the microorganism itself, but there are a few overarching mechanisms present. T. gondii oocysts have been found to resist an acidic pH and, thus, are protected by the acidification found in endocytic vacuoles and lysosomes.[76] Phagocytosis further increases with the carbohydrate-rich surface membrane located on the amoebae.[77] The pathogen can be released either by lysis of the amoebae or by exocytosis, but this is understudied [78]
Impact on wild birds
Almost all species of birds that have been tested for Toxoplasma gondii has shown to be positive. The only bird species not reported with clinical symptoms of toxoplasmosis would be wild ducks, and there has only been one report found on domesticated ducks occurring in 1962.[79] Species with resistance toward T. gondii include domestic turkeys,[80] owls, red tail hawks, and sparrows, depending on the strain of T. gondii infected.[81] T. gondii is considerably more severe in pigeons, particularly crown pigeons, ornamental pigeons, and pigeons originating from Australia and New Zealand. Typical onset is quick and usually is resulting in death. Those that do survive often have chronic conditions of encephalitis and neuritis.[81] Similarly, canaries are observed to be just as severe as pigeons, but the clinical symptoms are more abnormal when compared to other species. Most of the infection affects the eye, causing blindness, choroidal lesions, conjunctivitis, atrophy of the eye, blepharitis, and chorioretinitis [81] Most of the time, the infection leads to death.
Current environmental efforts
Urbanization and global warming are extremely influential in the transmission of T. gondii.[82] Temperature and humidity are huge factors in the sporulation stage: low humidity is always fatal to the oocysts, and they are also vulnerable to extreme temperatures.[82] Rainfall is also an important factor for survival of waterborne pathogens. Because increased rainfall directly increases the flow rate in rivers, the amount of flow into coastal areas is increased as well. This can spread waterborne pathogens over wide areas.
There is no effective vaccine for T. gondii, and research on a live vaccine is ongoing. Feeding cats commercially available food, rather than raw, undercooked meat, prevent felines from becoming a host for oocysts, as higher prevalence is in areas where raw meat is fed.[83] Researchers also suggest that owners restrict cats to live indoors and to be neutered or spayed to decrease stray cat populations and to reduce intermediate host interactions. Fecal matter from litter boxes is suggested to be collected daily, placed in a sealable bag, and disposed of in the trash and not flushed in the toilet, so that water contamination is limited.[84]
Studies have found that wetlands with a high density of vegetation decrease the concentration of oocysts in water through two possible mechanisms. Firstly, vegetation decreases flow velocities, which enables more settling because of increased transport time.[85] Secondly, the vegetation can remove oocysts through its ability to mechanically strain the water, as well as through the process of adhesion (i.e. attachment to biofilms). Areas of erosion and destruction of coastal wetlands have been found to harbour increased concentrations of T. gondii oocysts, which then flow into open coastal waters. Current physical and chemical treatments typically utilized in water treatment facilities have been proven to be ineffective against T. gondii. Research has shown that UV-C disinfection of water containing oocysts results in inactivation and possible sterilization.[86]
Genome
The genomes of more than 60 strains of T. gondii have been sequenced. Most are 60–80 Mb in size and consist of 11–14 chromosomes.[87][88] The major strains encode 7800–10,000 proteins, of which about 5200 are conserved across RH, GT1, ME49, VEG.[87] A database, ToxoDB, has been established to document genomic information on Toxoplasma.[89][90][91]
History
In 1908, while working at the Pasteur Institute in Tunis, Charles Nicolle and Louis Manceaux discovered a protozoan organism in the tissues of a hamster-like rodent known as the gundi, Ctenodactylus gundi.[30] Although Nicolle and Manceaux initially believed the organism to be a member of the genus Leishmania that they described as "Leishmania gondii", they soon realized they had discovered a new organism entirely; they renamed it Toxoplasma gondii. The new genus name Toxoplasma is a reference to its morphology: Toxo, from Greek τόξον (toxon, "arc, bow"), and πλάσμα (plasma, "shape, form") and the host in which it was discovered, the gundi (gondii).[92] The same year Nicolle and Mancaeux discovered T. gondii, Alfonso Splendore identified the same organism in a rabbit in Brazil. However, he did not give it a name.[30]
The first conclusive identification of T. gondii in humans was in an infant girl delivered full term by Caesarean section on May 23, 1938, at Babies' Hospital in New York City.[30] The girl began having seizures at three days of age, and doctors identified lesions in the maculae of both of her eyes. When she died at one month of age, an autopsy was performed. Lesions discovered in her brain and eye tissue were found to have both free and intracellular T. gondii'.[30] Infected tissue from the girl was homogenized and inoculated intracerebrally into rabbits and mice; they then developed encephalitis. Later, congenital transmission was confirmed in many other species, particularly infected sheep and rodents.
The possibility of T. gondii transmission via consumption of undercooked meat was first proposed by D. Weinman and A.H Chandler in 1954.[30] In 1960, the relevant cyst wall were shown to dissolve in the proteolytic enzymes found in the stomach, releasing infectious bradyzoites into the stomach (which pass into the intestine). The hypothesis of transmission via consumption of undercooked meat was tested in an orphanage in Paris in 1965; incidence of T. gondii rose from 10% to 50% after a year of adding two portions of cooked-rare beef or horse meat to many orphans' daily diets, and to 100% among those fed cooked-rare lamb chops.[30]
A 1959 Mumbai-based study found there prevalence in strict vegetarians was similar to that of non-vegetarians. This raised the possibility of a third major route of infection, beyond congenital and non well-cooked meat carnivorous transmission.[30]
In 1970, oocysts were found in (cat) feces. The fecal-oral route of infection via oocysts was demonstrated.[30] In the 1970s and 1980s feces of a vast range of infected animal species was tested to see if it contained oocysts — at least 17 species of felids shed oocysts, but no non-felid has been shown to allow T. gondii sexual reproduction (leading to oocyst shedding).[30]
Behavioral differences of infected hosts
There are many instances where behavioural changes were reported in rodents with T. gondii. The changes seen were a reduction in their innate dislike of cats, which made it easier for cats to prey on the rodents. In an experiment conducted by Berdoy and colleagues, the infected rats showed preference for the cat odour area versus the area with the rabbit scent, therefore making it easier for the parasite to take its final step in its definitive feline host.[11] This is an example of the extended phenotype concept, that is, the idea that the behaviour of the infected animal changes in order to maximize survival of the genes that increase predation of the intermediate rodent host.[93]
Differences in sex-dependent behavior observed in infected hosts compared to non-infected individuals can be attributed to differences in testosterone. Infected males had higher levels of testosterone while infected females had significantly lower levels, compared to their non-infected equivalents.[94] Looking at humans, studies using the Cattell's 16 Personality Factor questionnaire found that infected men scored lower on Factor G (superego strength/rule consciousness) and higher on Factor L (vigilance) while the opposite pattern was observed for infected women.[95] Such men were more likely to disregard rules and were more expedient, suspicious and jealous. On the other hand, women were more warm-hearted, outgoing, conscientious and moralistic.[95] Mice infected with T. gondii have a worse motor performance than non-infected mice.[96][97] Thus, a computerized simple reaction test was given to both infected and non-infected adults. It was found that the infected adults performed much more poorly and lost their concentration more quickly than the control group. But, the effect of the infection only explains less than 10% of the variability in performance[95] (i.e., there could be other confounding factors). Correlation has also been observed between seroprevalence of T. gondii in humans and increased risk of traffic accidents. Infected subjects have a 2.65 times higher risk of getting into a traffic accident.[98] A Turkish study confirmed this holds true among drivers.[99] This parasite has been associated with many neurological disorders such as schizophrenia. In a meta-analysis of 23 studies that met inclusion criteria, the seroprevalence of antibodies to T. gondii in people with schizophrenia is significantly higher than in control populations (OR=2.73, P<0.000001).[100] A 2009 summary of studies found that suicide attempters had far more indicative (IgG) antibodies than mental health inpatients without a suicide attempt.[101] Infection was also shown to be associated with suicide in women over the age of 60. (P<0.005) [102]
As mentioned before, these results of increased proportions of people seropositive for the parasite in cases of these neurological disorders do not necessarily indicate a causal relationship between the infection and disorder. It is also important to mention that in 2016 a population-representative birth cohort study which was done, to test a hypothesis that toxoplasmosis is related to impairment in brain and behaviour measured by a range of phenotypes including neuropsychiatric disorders, poor impulse control, personality and neurocognitive deficits. The results of this study did not support the results in the previously mentioned studies, more than marginally. None of the P-values showed significance for any outcome measure. Thus, according to this study, the presence of T. gondii antibodies is not correlated to increase susceptibility to any of the behaviour phenotypes (except possibly to a higher rate of unsuccessful attempted suicide). This team did not observe any significant association between T. gondii seropositivity and schizophrenia. The team notes that the null findings might be a false negative due to low statistical power because of small sample sizes, but against this weights that their set-up should avoid some possibilities for errors in the about 40 studies that did show a positive correlation. They concluded that further studies should be performed.[103] Another population-representative study with 7440 people in the United States found that Toxoplasma infection was 2.4 fold more common in people who had a history of manic and depression symptoms (bipolar disorder Type 1) compared to the general population.[104]
Research on the linkage between T. gondii infection and entrepreneurial behavior showed that students who tested positive for T. gondii exposure were 1.4 times more likely to major in business, and 1.7 times more likely to have an emphasis in "management and entrepreneurship". Among 197 participants of entrepreneurship events, T. gondii exposure was correlated with being 1.8 times more likely to have started their own business.[105]
The mechanism behind behavioral changes is partially attributed to increased dopamine metabolism,[106] which can be neutralized by dopamine antagonist medications.[107] T. gondii has two genes that code for a bifunctional phenylalanine and tyrosine hydroxylase, two important and rate-limiting steps of dopamine biosynthesis. One of the genes is constitutively expressed, while the other is only produced during cyst development.[108][109] In addition to additional dopamine production, T. gondii infection also produces long-lasting epigenetic changes in animals that increase the expression of vasopressin, a probable cause of alternations that persist after the clearance of the infection.[110]
References
- Nicolle, C.; Manceaux, L. (1909). "Sur un Protozoaire nouveau du Gondi". Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences. 148 (1): 369–72.
- Nicolle, C.; Manceaux, L. (1908). "Sur une infection à corps de Leishman (ou organismes voisins) du Gondi". Comptes Rendus Hebdomadaires des Séances de l'Académie des Sciences. 147 (2): 763–66.
- Dardé, M. L.; Ajzenberg, D.; Smith, J. (2011). "Population structure and epidemiology of Toxoplasma gondii". In Weiss, L. M.; Kim, K. (eds.). Toxoplasma Gondii: The Model Apicomplexan. Perspectives and Methods. Amsterdam, Boston, Heidelberg, London, New York: Elsevier. pp. 49–80. doi:10.1016/B978-012369542-0/50005-2. ISBN 978-0-12-369542-0.
- Dubey, J. P. (2010). "General Biology". Toxoplasmosis of Animals and Humans (Second ed.). Boca Raton, London, New York: Taylor and Francis Group. pp. 1–20. ISBN 978-1-4200-9237-0. Retrieved 1 February 2019.
- "CDC - Toxoplasmosis - Biology". 17 March 2015. Retrieved 14 June 2015.
- Knoll, Laura J.; Dubey, J. P.; Wilson, Sarah K.; Genova, Bruno Martorelli Di (2019-07-01). "Intestinal delta-6-desaturase activity determines host range for Toxoplasma sexual reproduction". bioRxiv: 688580. doi:10.1101/688580.
- "Cat parasite linked to mental illness, schizophrenia". CBS. Retrieved 23 September 2015.
- "CDC – About Parasites". Retrieved 12 March 2013.
- Flegr J, Prandota J, Sovičková M, Israili ZH (March 2014). "Toxoplasmosis – a global threat. Correlation of latent toxoplasmosis with specific disease burden in a set of 88 countries". PLOS ONE. 9 (3): e90203. Bibcode:2014PLoSO...990203F. doi:10.1371/journal.pone.0090203. PMC 3963851. PMID 24662942.
Toxoplasmosis is becoming a global health hazard as it infects 30–50% of the world human population. Clinically, the life-long presence of the parasite in tissues of a majority of infected individuals is usually considered asymptomatic. However, a number of studies show that this 'asymptomatic infection' may also lead to development of other human pathologies. ... The seroprevalence of toxoplasmosis correlated with various disease burden. Statistical associations does not necessarily mean causality. The precautionary principle suggests, however, that possible role of toxoplasmosis as a triggering factor responsible for development of several clinical entities deserves much more attention and financial support both in everyday medical practice and future clinical research.
- Pappas G, Roussos N, Falagas ME (October 2009). "Toxoplasmosis snapshots: global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis". International Journal for Parasitology. 39 (12): 1385–94. doi:10.1016/j.ijpara.2009.04.003. PMID 19433092.
- Berdoy M, Webster JP, Macdonald DW (August 2000). "Fatal attraction in rats infected with Toxoplasma gondii". Proceedings of the Royal Society of London B: Biological Sciences. 267 (1452): 1591–94. doi:10.1098/rspb.2000.1182. PMC 1690701. PMID 11007336.
- "CDC Parasites – Toxoplasmosis (Toxoplasma infection) – Disease". Retrieved 12 March 2013.
- Cook TB, Brenner LA, Cloninger CR, Langenberg P, Igbide A, Giegling I, Hartmann AM, Konte B, Friedl M, Brundin L, Groer MW, Can A, Rujescu D, Postolache TT (January 2015). ""Latent" infection with Toxoplasma gondii: association with trait aggression and impulsivity in healthy adults". Journal of Psychiatric Research. 60: 87–94. doi:10.1016/j.jpsychires.2014.09.019. PMID 25306262.
- Karen Sugden , Terrie E. Moffitt, Lauriane Pinto, Richie Poulton, Benjamin S. Williams, Avshalom Caspi (17 February 2016). "Is Toxoplasma Gondii Infection Related to Brain and Behavior Impairments in Humans? Evidence from a Population-Representative Birth Cohort". PLOS ONE. 11 (2): e0148435. Bibcode:2016PLoSO..1148435S. doi:10.1371/journal.pone.0148435. PMC 4757034. PMID 26886853.CS1 maint: uses authors parameter (link)
- Webster JP (May 2007). "The effect of Toxoplasma gondii on animal behavior: playing cat and mouse". Schizophrenia Bulletin. 33 (3): 752–6. doi:10.1093/schbul/sbl073. PMC 2526137. PMID 17218613.
- Webster JP, Kaushik M, Bristow GC, McConkey GA (January 2013). "Toxoplasma gondii infection, from predation to schizophrenia: can animal behaviour help us understand human behaviour?". The Journal of Experimental Biology. 216 (Pt 1): 99–112. doi:10.1242/jeb.074716. PMC 3515034. PMID 23225872.
- Hari Dass SA, Vyas A (December 2014). "Toxoplasma gondii infection reduces predator aversion in rats through epigenetic modulation in the host medial amygdala". Molecular Ecology. 23 (24): 6114–22. doi:10.1111/mec.12888. PMID 25142402.
- Flegr J, Markoš A (December 2014). "Masterpiece of epigenetic engineering – how Toxoplasma gondii reprogrammes host brains to change fear to sexual attraction". Molecular Ecology. 23 (24): 5934–36. doi:10.1111/mec.13006. PMID 25532868.
- Vanagas L, Jeffers V, Bogado SS, Dalmasso MC, Sullivan WJ, Angel SO (October 2012). "Toxoplasma histone acetylation remodelers as novel drug targets". Expert Review of Anti-Infective Therapy. 10 (10): 1189–201. doi:10.1586/eri.12.100. PMC 3581047. PMID 23199404.
- Bouchut A, Chawla AR, Jeffers V, Hudmon A, Sullivan WJ (2015). "Proteome-wide lysine acetylation in cortical astrocytes and alterations that occur during infection with brain parasite Toxoplasma gondii". PLOS ONE. 10 (3): e0117966. Bibcode:2015PLoSO..1017966B. doi:10.1371/journal.pone.0117966. PMC 4364782. PMID 25786129.
- Flegr J, Lenochová P, Hodný Z, Vondrová M (November 2011). "Fatal attraction phenomenon in humans: cat odour attractiveness increased for toxoplasma-infected men while decreased for infected women". PLOS Neglected Tropical Diseases. 5 (11): e1389. doi:10.1371/journal.pntd.0001389. PMC 3210761. PMID 22087345.
- Flegr J (January 2013). "Influence of latent Toxoplasma infection on human personality, physiology and morphology: pros and cons of the Toxoplasma-human model in studying the manipulation hypothesis". The Journal of Experimental Biology. 216 (Pt 1): 127–33. doi:10.1242/jeb.073635. PMID 23225875.
- Pearce BD, Kruszon-Moran D, Jones JL (2012). "The Relationship Between Toxoplasma Gondii Infection and Mood Disorders in the Third National Health and Nutrition Survey". Biological Psychiatry. 72 (4): 290–95. doi:10.1016/j.biopsych.2012.01.003. PMC 4750371. PMID 22325983.
- de Barros JL, Barbosa IG, Salem H, Rocha NP, Kummer A, Okusaga OO, Soares JC, Teixeira AL (February 2017). "Is there any association between Toxoplasma gondii infection and bipolar disorder? A systematic review and meta-analysis". Journal of Affective Disorders. 209: 59–65. doi:10.1016/j.jad.2016.11.016. PMID 27889597.
- Gale SD, Erickson LD, Brown BL, Hedges DW (2015). "Interaction between Helicobacter pylori and latent toxoplasmosis and demographic variables on cognitive function in young to middle-aged adults". PLOS ONE. 10 (1): e0116874. Bibcode:2015PLoSO..1016874G. doi:10.1371/journal.pone.0116874. PMC 4295891. PMID 25590622.
- Parlog A, Schlüter D, Dunay IR (March 2015). "Toxoplasma gondii-induced neuronal alterations". Parasite Immunology. 37 (3): 159–70. doi:10.1111/pim.12157. hdl:10033/346575. PMID 25376390.
- Blanchard N, Dunay IR, Schlüter D (March 2015). "Persistence of Toxoplasma gondii in the central nervous system: a fine-tuned balance between the parasite, the brain and the immune system". Parasite Immunology. 37 (3): 150–58. doi:10.1111/pim.12173. hdl:10033/346515. PMID 25573476.
- Weiss LM, Kim K (2011). Toxoplasma Gondii: The Model Apicomplexan. Perspectives and Methods. Academic Press. ISBN 978-0-08-047501-1. Retrieved 12 March 2013.
- Dubey JP, Ferreira LR, Martins J, Jones JL (October 2011). "Sporulation and survival of Toxoplasma gondii oocysts in different types of commercial cat litter". The Journal of Parasitology. 97 (5): 751–54. doi:10.1645/GE-2774.1. PMID 21539466. S2CID 41292680.
- Dubey JP (July 2009). "History of the discovery of the life cycle of Toxoplasma gondii". International Journal for Parasitology. 39 (8): 877–82. doi:10.1016/j.ijpara.2009.01.005. PMID 19630138.
- Kapperud G, Jenum PA, Stray-Pedersen B, Melby KK, Eskild A, Eng J (August 1996). "Risk factors for Toxoplasma gondii infection in pregnancy. Results of a prospective case-control study in Norway". American Journal of Epidemiology. 144 (4): 405–12. doi:10.1093/oxfordjournals.aje.a008942. PMID 8712198.
- Dubey JP (July 1998). "Advances in the life cycle of Toxoplasma gondii". International Journal for Parasitology. 28 (7): 1019–24. doi:10.1016/S0020-7519(98)00023-X. PMID 9724872.
- Robert-Gangneux F, Dardé ML (April 2012). "Epidemiology of and diagnostic strategies for toxoplasmosis". Clinical Microbiology Reviews. 25 (2): 264–96. doi:10.1128/CMR.05013-11. PMC 3346298. PMID 22491772.
- Markus, MB (1987). "Terms for coccidian merozoites". Annals of Tropical Medicine & Parasitology. 81 (4): 463. doi:10.1080/00034983.1987.11812147. PMID 3446034.
- Miller CM, Boulter NR, Ikin RJ, Smith NC (January 2009). "The immunobiology of the innate response to Toxoplasma gondii". International Journal for Parasitology. 39 (1): 23–39. doi:10.1016/j.ijpara.2008.08.002. PMID 18775432.
- "CDC Toxoplasmosis – Microscopy Findings". Retrieved 13 March 2013.
- Clarence R. Robbins (2012). Chemical and Physical Behavior of Human Hair. Springer. p. 585. ISBN 978-3-642-25610-3. Retrieved 12 March 2013.
- Jones JL, Dubey JP (September 2012). "Foodborne toxoplasmosis". Clinical Infectious Diseases. 55 (6): 845–51. doi:10.1093/cid/cis508. PMID 22618566.
- "Parasite Shed in Cat Feces Kills Sea Otters – California Sea Grant" (PDF). www-csgc.ucsd.edu.
- Khan A, Böhme U, Kelly KA, Adlem E, Brooks K, Simmonds M, Mungall K, Quail MA, Arrowsmith C, Chillingworth T, Churcher C, Harris D, Collins M, Fosker N, Fraser A, Hance Z, Jagels K, Moule S, Murphy L, O'Neil S, Rajandream MA, Saunders D, Seeger K, Whitehead S, Mayr T, Xuan X, Watanabe J, Suzuki Y, Wakaguri H, Sugano S, Sugimoto C, Paulsen I, Mackey AJ, Roos DS, Hall N, Berriman M, Barrell B, Sibley LD, Ajioka JW (September 2006). "Common inheritance of chromosome Ia associated with clonal expansion of Toxoplasma gondii". Genome Research. 16 (9): 1119–25. doi:10.1101/gr.5318106. PMC 1557770. PMID 16902086.
- Dubey JP, Velmurugan GV, Rajendran C, Yabsley MJ, Thomas NJ, Beckmen KB, Sinnett D, Ruid D, Hart J, Fair PA, McFee WE, Shearn-Bochsler V, Kwok OC, Ferreira LR, Choudhary S, Faria EB, Zhou H, Felix TA, Su C (September 2011). "Genetic characterisation of Toxoplasma gondii in wildlife from North America revealed widespread and high prevalence of the fourth clonal type". International Journal for Parasitology. 41 (11): 1139–47. doi:10.1016/j.ijpara.2011.06.005. PMID 21802422.
- Rigoulet J, Hennache A, Lagourette P, George C, Longeart L, Le Net JL, Dubey JP (2014). "Toxoplasmosis in a bar-shouldered dove (Geopelia humeralis) from the Zoo of Clères, France". Parasite. 21: 62. doi:10.1051/parasite/2014062. PMC 4236686. PMID 25407506.

- Parks S, Avramopoulos D, Mulle J, McGrath J, Wang R, Goes F S, Conneely K, Ruczinski I, Yolken R, Pulver A E. HLA typing using genome wide data reveals susceptibility types for infections in a psychiatric disease enriched sample. Brain, Behav, Immun. 2018.
- Henriquez SA, Brett R, Alexander J, Pratt J, Roberts CW (2009). "Neuropsychiatric disease and Toxoplasma gondii infection". Neuroimmunomodulation. 16 (2): 122–33. doi:10.1159/000180267. PMID 19212132.
- Konsman JP, Parnet P, Dantzer R (March 2002). "Cytokine-induced sickness behaviour: mechanisms and implications". Trends in Neurosciences. 25 (3): 154–59. doi:10.1016/s0166-2236(00)02088-9. PMID 11852148.
- Tenter AM, Heckeroth AR, Weiss LM (November 2000). "Toxoplasma gondii: from animals to humans". International Journal for Parasitology. 30 (12–13): 1217–58. doi:10.1016/S0020-7519(00)00124-7. PMC 3109627. PMID 11113252.
- Jones JL, Dargelas V, Roberts J, Press C, Remington JS, Montoya JG (September 2009). "Risk factors for Toxoplasma gondii infection in the United States". Clinical Infectious Diseases. 49 (6): 878–84. doi:10.1086/605433. PMID 19663709.
- Cook AJ, Gilbert RE, Buffolano W, Zufferey J, Petersen E, Jenum PA, Foulon W, Semprini AE, Dunn DT (July 2000). "Sources of toxoplasma infection in pregnant women: European multicentre case-control study. European Research Network on Congenital Toxoplasmosis". BMJ. 321 (7254): 142–47. doi:10.1136/bmj.321.7254.142. PMC 27431. PMID 10894691.
- Sakikawa M, Noda S, Hanaoka M, Nakayama H, Hojo S, Kakinoki S, Nakata M, Yasuda T, Ikenoue T, Kojima T (March 2012). "Anti-Toxoplasma antibody prevalence, primary infection rate, and risk factors in a study of toxoplasmosis in 4,466 pregnant women in Japan". Clinical and Vaccine Immunology. 19 (3): 365–67. doi:10.1128/CVI.05486-11. PMC 3294603. PMID 22205659.
- Dubey JP, Hill DE, Jones JL, Hightower AW, Kirkland E, Roberts JM, Marcet PL, Lehmann T, Vianna MC, Miska K, Sreekumar C, Kwok OC, Shen SK, Gamble HR (October 2005). "Prevalence of viable Toxoplasma gondii in beef, chicken, and pork from retail meat stores in the United States: risk assessment to consumers". The Journal of Parasitology. 91 (5): 1082–93. doi:10.1645/ge-683.1. PMID 16419752.
- Mai K, Sharman PA, Walker RA, Katrib M, De Souza D, McConville MJ, Wallach MG, Belli SI, Ferguson DJ, Smith NC (March 2009). "Oocyst wall formation and composition in coccidian parasites". Memórias do Instituto Oswaldo Cruz. 104 (2): 281–89. doi:10.1590/S0074-02762009000200022. PMID 19430654.
- Siegel SE, Lunde MN, Gelderman AH, Halterman RH, Brown JA, Levine AS, Graw RG (April 1971). "Transmission of toxoplasmosis by leukocyte transfusion". Blood. 37 (4): 388–94. doi:10.1182/blood.V37.4.388.388. PMID 4927414.
- Gallas-Lindemann C, Sotiriadou I, Mahmoodi MR, Karanis P (February 2013). "Detection of Toxoplasma gondii oocysts in different water resources by Loop Mediated Isothermal Amplification (LAMP)". Acta Tropica. 125 (2): 231–36. doi:10.1016/j.actatropica.2012.10.007. PMID 23088835.
- Alvarado-Esquivel C, Liesenfeld O, Márquez-Conde JA, Estrada-Martínez S, Dubey JP (October 2010). "Seroepidemiology of infection with Toxoplasma gondii in workers occupationally exposed to water, sewage, and soil in Durango, Mexico". The Journal of Parasitology. 96 (5): 847–50. doi:10.1645/GE-2453.1. PMID 20950091. S2CID 23241017.
- Esmerini PO, Gennari SM, Pena HF (May 2010). "Analysis of marine bivalve shellfish from the fish market in Santos city, São Paulo state, Brazil, for Toxoplasma gondii". Veterinary Parasitology. 170 (1–2): 8–13. doi:10.1016/j.vetpar.2010.01.036. PMID 20197214.
- Dattoli VC, Veiga RV, Cunha SS, Pontes-de-Carvalho L, Barreto ML, Alcantara-Neves NM (December 2011). "Oocyst ingestion as an important transmission route of Toxoplasma gondii in Brazilian urban children". The Journal of Parasitology. 97 (6): 1080–84. doi:10.1645/GE-2836.1. PMID 21740247.
- Bobić B, Jevremović I, Marinković J, Sibalić D, Djurković-Djaković O (September 1998). "Risk factors for Toxoplasma infection in a reproductive age female population in the area of Belgrade, Yugoslavia". European Journal of Epidemiology. 14 (6): 605–10. doi:10.1023/A:1007461225944. PMID 9794128. S2CID 9423818.
- Jones JL, Dargelas V, Roberts J, Press C, Remington JS, Montoya JG (September 2009). "Risk factors for Toxoplasma gondii infection in the United States". Clinical Infectious Diseases. 49 (6): 878–84. doi:10.1086/605433. PMID 19663709.
- Dass SA, Vasudevan A, Dutta D, Soh LJ, Sapolsky RM, Vyas A (2011). "Protozoan parasite Toxoplasma gondii manipulates mate choice in rats by enhancing attractiveness of males". PLOS ONE. 6 (11): e27229. Bibcode:2011PLoSO...627229D. doi:10.1371/journal.pone.0027229. PMC 3206931. PMID 22073295.
- Arantes TP, Lopes WD, Ferreira RM, Pieroni JS, Pinto VM, Sakamoto CA, Costa AJ (October 2009). "Toxoplasma gondii: Evidence for the transmission by semen in dogs". Experimental Parasitology. 123 (2): 190–94. doi:10.1016/j.exppara.2009.07.003. PMID 19622353.
- Gutierrez J, O'Donovan J, Williams E, Proctor A, Brady C, Marques PX, Worrall S, Nally JE, McElroy M, Bassett H, Sammin D, Buxton D, Maley S, Markey BK (August 2010). "Detection and quantification of Toxoplasma gondii in ovine maternal and foetal tissues from experimentally infected pregnant ewes using real-time PCR". Veterinary Parasitology. 172 (1–2): 8–15. doi:10.1016/j.vetpar.2010.04.035. PMID 20510517.
- "CDC: Parasites – Toxoplasmosis (Toxoplasma infection) – Prevention & Control". Retrieved 13 March 2013.
- "Mayo Clinic – Toxoplasmosis – Prevention". Retrieved 13 March 2013.
- Green, Aliza (2005). Field Guide to Meat. Philadelphia, PA: Quirk Books. pp. 294–95. ISBN 978-1-59474-017-6.
- Verma R, Khanna P (February 2013). "Development of Toxoplasma gondii vaccine: A global challenge". Human Vaccines & Immunotherapeutics. 9 (2): 291–93. doi:10.4161/hv.22474. PMC 3859749. PMID 23111123.
- "TOXPOX Result In Brief – Vaccine against Toxoplasmosis". CORDIS, European Commission. 2015-01-14. Retrieved 2015-12-11.
- "TOXOVAX®". MSD Animal Health. Retrieved 2015-11-10.
- Hollings, T., Jones, M., Mooney, N., & McCallum, H. (2013). Wildlife disease ecology in changing landscapes: Mesopredator release and toxoplasmosis. International Journal for Parasitology: Parasites and Wildlife, 110–18.
- Conrad, P., Miller, M., Kreuder, C., James, E., Mazer, J., Dabritz, H., ... Gulland, F. G. (2005). Transmission of Toxoplasma: Clues from the study of sea otters as sentinels of toxoplasma gondii flow into the marine environment. International Journal for Parasitology , 1155–68.
- Defenders of Wildlife. (2020). Sea Otter. Retrieved from Defenders of Wildlife: https://defenders.org/wildlife/sea-otter
- Ahlers, A. A., A., M. M., Dubey, J. P., & Schooley, R. L. (2015). Risk Factors for Toxoplasma gondii Exposure in Semiaquatic Mammals in a Freshwater Ecosystem. Wildlife Diseases, 488–92.
- I.C.L., A., A.F., S.-F., Munoz-Leal, S., Soares, H., M.B., H., Moreno, L., ... Gennari, S. (2019). Evaluation of antibodies against Toxoplasma gondii and Leptospira spp. in Magellanic penguins (Speniscus magellanicus) on Magdalena Island, Chile. Veterinary Parasitology: Regional Studies and Reports, 1–4.
- Ploeg, M., Ultee, T., & Kik, M. (2011). Disseminated Toxoplasmosis in Black-Footed Penguins (Spheniscus demersus). Avian Diseases, 55(4), 701–03.
- Greub, G., & Raoult, D. (2004). Microorganisms Resistant to Free-Living Amoebae. Clinical Microbiology Reviews, 413–33.
- Cirillo, J., Falkow, S., Tompkins, L., & Bermundez, L. (1997). Interaction of Mycobacterium avium with environmental amoebae enhances virulence. Infection and Immunity, 3759–67.
- Winiecka-Krusnell, J., Dellacasa-Lindberg, I., Dubey, J., & Barragan, A. (2009). Toxoplasma gondii: Uptake and survival of oocysts in free-living amoebae. Experimental Parasitology, 124–31.
- Elloway, E., Armstrong, R., Bird, R., Kelly, S., & Smith, S. (2004). Analysis of Acanthamoebapolyphaga surface carbohydrate exposure by FITC-lectin binding and fluorescence evaluation. Journal of Applied Microbiology, 1319–25.
- Paquet, V. E., & Charette, S. J. (2016). Amoeba-resisting bacteria found in multilamellar bodies secreted by Dictyostelium discoideum: social amoebae can also package bacteria. Microbiology Ecology , 1–8.
- Boehringer, E. G., Fornari, O. E., & Boehringer, I. K. (1962). The first case of toxoplasma gondii in domestic ducks in Argentina. Avian Diseases, 391–96.
- Drobeck, H. P., Manwell, R. D., Bernstein, E., & Dillon, R. D. (1953). Further studies of toxoplasmosis in birds. American Journal of Epidemiology, 329–39.
- Dubey, J. (2002). A review of toxoplasmosis in wild birds. Veterinary Parasitology, 121–53.
- Yan, C., Liang, L.-J., Zheng, K.-Y., & Zhu, X.-Q. (2016). Impact of environmental factors on the emrgence, transmission and distribution of Toxoplasma gondii. Parasites & Vectors, 1–7.
- Elmore, S. A., Jones, J. L., Conrad, P. A., Patton, S., Lindsay, D. S., & Dubey, J. (2010). Toxoplasma gondii: epidemiology, feline clinical aspects, and prevention. Trends in Parasitology, 190–96.
- Shapiro, K., Bahia-Oliveira, L., Dixon, B., Dumetre, A., de Wit, L. A., VanWormer, E., & Villena, i. (2019). Environmental transmission of toxoplasma gondii: Oocysts in water, soil and food. Food and Waterborne Parasitology, 1–18.
- Shapiro, K., Bahia-Oliveira, L., Dixon, B., Dumetre, A., de Wit, L. A., VanWormer, E., & Villena, i. (2019). Environmental tranmission of toxoplasma gondii: Oocysts in water, soil and food. Food and Waterborne Parasitology, 1-18.
- Dumetre, A., Le Bras, C., Baffet, M., Meneceur, P., Dubey, J., Derouin, F., ... Moulin, L. (2008). Effects of ozone and ultraviolet radiation treatments on the infectivity of toxoplasma gondii oocysts. Veterinary Parasitology, 209–13.
- Lau YL, Lee WC, Gudimella R, Zhang G, Ching XT, Razali R, Aziz F, Anwar A, Fong MY (2016-06-29). "Deciphering the Draft Genome of Toxoplasma gondii RH Strain". PLOS ONE. 11 (6): e0157901. Bibcode:2016PLoSO..1157901L. doi:10.1371/journal.pone.0157901. PMC 4927122. PMID 27355363.
- Bontell IL, Hall N, Ashelford KE, Dubey JP, Boyle JP, Lindh J, Smith JE (2009-05-20). "Whole genome sequencing of a natural recombinant Toxoplasma gondii strain reveals chromosome sorting and local allelic variants". Genome Biology. 10 (5): R53. doi:10.1186/gb-2009-10-5-r53. PMC 2718519. PMID 19457243.
- Kissinger JC, Gajria B, Li L, Paulsen IT, Roos DS (January 2003). "ToxoDB: accessing the Toxoplasma gondii genome". Nucleic Acids Research. 31 (1): 234–36. doi:10.1093/nar/gkg072. PMC 165519. PMID 12519989.
- Gajria B, Bahl A, Brestelli J, Dommer J, Fischer S, Gao X, Heiges M, Iodice J, Kissinger JC, Mackey AJ, Pinney DF, Roos DS, Stoeckert CJ, Wang H, Brunk BP (January 2008). "ToxoDB: an integrated Toxoplasma gondii database resource". Nucleic Acids Research. 36 (Database issue): D553–56. doi:10.1093/nar/gkm981. PMC 2238934. PMID 18003657.
- "ToxoDB : The Toxoplasma Genomics Resource". toxodb.org. Retrieved 2018-03-01.
- Flegr, Jaroslav; Prandota, Joseph; Sovičková, Michaela; Israili, Zafar H. (2014-03-24). "Toxoplasmosis – A Global Threat. Correlation of Latent Toxoplasmosis with Specific Disease Burden in a Set of 88 Countries". PLOS ONE. 9 (3): e90203. Bibcode:2014PLoSO...990203F. doi:10.1371/journal.pone.0090203. ISSN 1932-6203. PMC 3963851. PMID 24662942.
- McConkey GA, Martin HL, Bristow GC, Webster JP (January 2013). "Toxoplasma gondii infection and behaviour – location, location, location?". The Journal of Experimental Biology. 216 (Pt 1): 113–19. doi:10.1242/jeb.074153. PMC 3515035. PMID 23225873.
- Flegr J, Lindová J, Kodym P (April 2008). "Sex-dependent toxoplasmosis-associated differences in testosterone concentration in humans". Parasitology. 135 (4): 427–31. doi:10.1017/S0031182007004064. PMID 18205984.
- Flegr J (May 2007). "Effects of toxoplasma on human behavior". Schizophrenia Bulletin. 33 (3): 757–60. doi:10.1093/schbul/sbl074. PMC 2526142. PMID 17218612.
- Hrdá S, Votýpka J, Kodym P, Flegr J (August 2000). "Transient nature of Toxoplasma gondii-induced behavioral changes in mice". The Journal of Parasitology. 86 (4): 657–63. doi:10.1645/0022-3395(2000)086[0657:TNOTGI]2.0.CO;2. PMID 10958436.
- Hutchison WM, Aitken PP, Wells BW (October 1980). "Chronic Toxoplasma infections and motor performance in the mouse". Annals of Tropical Medicine and Parasitology. 74 (5): 507–10. doi:10.1080/00034983.1980.11687376. PMID 7469564.
- Flegr J, Havlícek J, Kodym P, Malý M, Smahel Z (July 2002). "Increased risk of traffic accidents in subjects with latent toxoplasmosis: a retrospective case-control study". BMC Infectious Diseases. 2: 11. doi:10.1186/1471-2334-2-11. PMC 117239. PMID 12095427.
- Kocazeybek B, Oner YA, Turksoy R, Babur C, Cakan H, Sahip N, Unal A, Ozaslan A, Kilic S, Saribas S, Aslan M, Taylan A, Koc S, Dirican A, Uner HB, Oz V, Ertekin C, Kucukbasmaci O, Torun MM (May 2009). "Higher prevalence of toxoplasmosis in victims of traffic accidents suggest increased risk of traffic accident in Toxoplasma-infected inhabitants of Istanbul and its suburbs". Forensic Science International. 187 (1–3): 103–08. doi:10.1016/j.forsciint.2009.03.007. PMID 19356869.
- Torrey EF, Bartko JJ, Lun ZR, Yolken RH (May 2007). "Antibodies to Toxoplasma gondii in patients with schizophrenia: a meta-analysis". Schizophrenia Bulletin. 33 (3): 729–36. doi:10.1093/schbul/sbl050. PMC 2526143. PMID 17085743.
- Arling TA, Yolken RH, Lapidus M, Langenberg P, Dickerson FB, Zimmerman SA, Balis T, Cabassa JA, Scrandis DA, Tonelli LH, Postolache TT (December 2009). "Toxoplasma gondii antibody titers and history of suicide attempts in patients with recurrent mood disorders". The Journal of Nervous and Mental Disease. 197 (12): 905–08. doi:10.1097/nmd.0b013e3181c29a23. PMID 20010026. S2CID 33395780.
- Ling VJ, Lester D, Mortensen PB, Langenberg PW, Postolache TT (July 2011). "Toxoplasma gondii seropositivity and suicide rates in women". The Journal of Nervous and Mental Disease. 199 (7): 440–44. doi:10.1097/nmd.0b013e318221416e. PMC 3128543. PMID 21716055.
- Sugden K, Moffitt TE, Pinto L, Poulton R, Williams BS, Caspi A (2016). "Is Toxoplasma Gondii Infection Related to Brain and Behavior Impairments in Humans? Evidence from a Population-Representative Birth Cohort". PLOS ONE. 11 (2): e0148435. Bibcode:2016PLoSO..1148435S. doi:10.1371/journal.pone.0148435. PMC 4757034. PMID 26886853.
- Pearce, B. D.; Kruszon-Moran, D.; Jones, J. L. (2012). "The Relationship Between Toxoplasma Gondii Infection and Mood Disorders in the Third National Health and Nutrition Survey". Biological Psychiatry. 72 (4): 290–95. doi:10.1016/j.biopsych.2012.01.003. PMC 4750371. PMID 22325983.
- Johnson, S. K.; Fitza, M. A.; Lerner, D. A.; Calhoun, D. M.; Beldon, M. A.; Chan, E. T.; Johnson, P. T. (2018). "Risky business: linking Toxoplasma gondii infection and entrepreneurship behaviours across individuals and countries". Proceedings of the Royal Society B: Biological Sciences. 285 (1883): 20180822. doi:10.1098/rspb.2018.0822. PMC 6083268. PMID 30051870.
- Prandovszky, E; Gaskell, E; Martin, H; Dubey, JP; Webster, JP; McConkey, GA (2011). "The neurotropic parasite Toxoplasma gondii increases dopamine metabolism". PLOS ONE. 6 (9): e23866. Bibcode:2011PLoSO...623866P. doi:10.1371/journal.pone.0023866. PMC 3177840. PMID 21957440.
- Webster, JP; Lamberton, PH; Donnelly, CA; Torrey, EF (22 April 2006). "Parasites as causative agents of human affective disorders? The impact of anti-psychotic, mood-stabilizer and anti-parasite medication on Toxoplasma gondii's ability to alter host behaviour". Proceedings. Biological Sciences. 273 (1589): 1023–30. doi:10.1098/rspb.2005.3413. PMC 1560245. PMID 16627289.
- Gaskell, EA; Smith, JE; Pinney, JW; Westhead, DR; McConkey, GA (2009). "A unique dual activity amino acid hydroxylase in Toxoplasma gondii". PLOS ONE. 4 (3): e4801. Bibcode:2009PLoSO...4.4801G. doi:10.1371/journal.pone.0004801. PMC 2653193. PMID 19277211.
- Sangrador, Amaia; Mitchell, Alex (6 November 2014). "Protein focus: Don't blame the cat – the toxoplasmosis effect". InterPro database blog. Retrieved 27 May 2019.
- Hari Dass, SA; Vyas, A (December 2014). "Toxoplasma gondii infection reduces predator aversion in rats through epigenetic modulation in the host medial amygdala". Molecular Ecology. 23 (24): 6114–22. doi:10.1111/mec.12888. PMID 25142402.
External links
- ToxoDB : The Toxoplasma gondii genome resource
- Anti-Toxo : A Toxoplasma news blog and list of research laboratories
- Toxoplasma images, from CDC's DPDx, in the public domain
- Toxoplasmosis Research Institute & Center
- Toxoplasma gondii seroprevalence varies by cat breed PLOS One 2017
- Neurological and Neuropsychiatric Consequences of Chronic Toxoplasma Infection ISSN 2196-5471, 2015
- Cytoskeletal Components of an Invasion Machine – The Apical Complex of Toxoplasma gondii
- The Culture-Shaping Parasites, in Seed Magazine
- Sneaky Parasite Attracts Rats to Cats, All Things Considered, April 14, 2007
- Toxoplasma lecture, Robert Sapolsky
- Could a brain parasite found in cats help soccer teams win at the World Cup?, – By Patrick House – Slate Magazine
- How Your Cat Is Making You Crazy, the Atlantic Magazine, March 2012
- Mystery Marine Mammal Deaths, CosmosMagazine.com, June 2008
- Toxoplasma gondii in the Subarctic and Arctic
- Okusaga O, Postolache TT (2012). "19. Toxoplasma gondii, the Immune System, and Suicidal Behaviour". In Dwivedi Y (ed.). The Neurobiological Basis of Suicide. CRC Press. ISBN 978-1-4398-3881-5. PMID 23035283.