Pseudopodia
A pseudopod or pseudopodium (plural: pseudopods or pseudopodia) is a temporary arm-like projection of a eukaryotic cell membrane that are developed in the direction of movement. Filled with cytoplasm, pseudopodia primarily consist of actin filaments and may also contain microtubules and intermediate filaments.[1][2] Pseudopods are used for motility and ingestion. They are often found in amoebas.
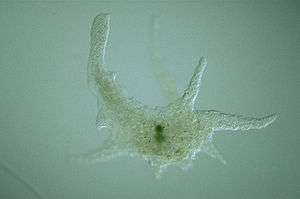
Different types of pseudopodia can be classified by their distinct appearances.[3] Lamellipodia are broad and thin. Filopodia are slender, thread-like, and are supported largely by microfilaments. Lobopodia are bulbous and amoebic. Reticulopodia are complex structures bearing individual pseudopodia which form irregular nets. Axopodia are the phagocytosis type with long, thin pseudopods supported by complex microtubule arrays enveloped with cytoplasm; they respond rapidly to physical contact.[4]
However some pseudopodial cells are able to use multiple types of pseudopodia depending on the situation: Most of them use a combination of lamellipodia and filopodia to migrate[5] (e.g. metastatic cancer cells[6]). The human foreskin fibroblasts can either use lamellipodia- or lobopodia-based migration in a 3D matrix depending on the matrix elasticity.[7]
Generally, several pseudopodia arise from the surface of the body, (polypodial, e.g. Amoeba proteus), or a single pseudopod may form on the surface of the body (monopodial, e.g. Entamoeba histolytica).[8]
Cells which make pseudopods are generally referred to as amoeboids.[9]
Formation
Via extracellular cue
To move towards a target, the cell uses chemotaxis. It senses extracellular signalling molecules, chemoattractants (e.g. cAMP for Dictyostelium cells[10]), to extend pseudopodia at the membrane area facing the source of these molecules.
The chemoattractants bind to G protein-coupled receptors, which activate GTPases of the Rho family (e.g. Cdc42, Rac) via G-proteins.
Rho GTPases are able to activate WASp which in turn activate Arp2/3 complex which serve as nucleation sites for actin polymerization.[11] The actin polymers then push the membrane as they grow, forming the pseudopod. The pseudopodium can then adhere to a surface via its adhesion proteins (e.g. integrins), and then pull the cell's body forward via contraction of an actin-myosin complex in the pseudopod.[12][13] This type of locomotion is called Amoeboid movement.
Rho GTPases can also activate phosphatidylinositol 3-kinase (PI3K) which recruit PIP3 to the membrane at the leading edge and detach the PIP3-degrading enzyme PTEN from the same area of the membrane. PIP3 then activate GTPases back via GEF stimulation. This serves as a feedback loop to amplify and maintain the presence of local GTPase at the leading edge.[11]
Otherwise, pseudopodia can't grow on other sides of the membrane than the leading edge because myosin filaments prevent them to extend. These myosin filaments are induced by cyclic GMP in D. discoideum or Rho kinase in neutrophils for example.[11]
Without extracellular cue
In the case there is no extracellular cue, all moving cells navigate in random directions, but they can keep the same direction for some time before turning. This feature allows cells to explore large areas for colonization or searching for a new extracellular cue.
In Dictyostelium cells, a pseudopodium can form either de novo as normal, or from an existing pseudopod, forming a Y-shaped pseudopodium.
The Y-shaped pseudopods are used by Dictyostelium to advance relatively straight forward by alternating between retraction of the left or right branch of the pseudopod. The de novo pseudopodia form at different sides than pre-existing ones, they are used by the cells to turn.
Y-shaped pseudopods are more frequent than de novo ones, which explain the preference of the cell to keep moving to the same direction. This persistence is modulated by PLA2 and cGMP signalling pathways.[10]
Functions
The functions of pseudopodia include locomotion and ingestion:
- Pseudopodia are critical in sensing targets which can then be engulfed; the engulfing pseudopodia are called phagocytosis pseudopodia. A common example of this type of amoeboid cell is the macrophage.
- They are also essential to amoeboid-like locomotion. Human mesenchymal stem cells are a good example of this function: these migratory cells are responsible for in-utero remodeling; for example, in the formation of the trilaminar germ disc during gastrulation.[14]
Morphology
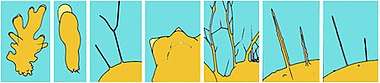
Pseudopods can be classified into several varieties according to the number of projections (monopodia and polypodia), and according to their appearance:
Lamellipodia
Lamellipodia are broad and flat pseudopodia used in locomotion.[4] They are supported by microfilaments which form at the leading edge, creating a mesh-like internal network.[15]
Filopodia
Filopodia (or filose pseudopods) are slender and filiform with pointed ends, consisting mainly of ectoplasm. These formations are supported by microfilaments which, unlike the filaments of lamellipodia with their net-like actin, form loose bundles by cross-linking. This formation is partly due to bundling proteins such as fimbrins and fascins.[15][16] Filopodia are observed in some animal cells: in part of Filosa (Rhizaria), in "Testaceafilosia" , in Vampyrellidae and Pseudosporida (Rhizaria) and in Nucleariida (Opisthokonta).[4]
Lobopodia
Lobopodia (or lobose pseudopods) are bulbous, short, and blunt in form.[17] These finger-like, tubular pseudopodia contain both ectoplasm and endoplasm. They can be found in different kind of cells, notably in Lobosa and other Amoebozoa and in some Heterolobosea (Excavata).
High-pressure lobopodia can also be found in human fibroblasts travelling through a complex network of 3D matrix (e.g. mammalian dermis, cell-derived matrix). Contrarily to other pseudopodia using the pressure exerted by actin polymerization on the membrane to extend, fibroblast lobopods use the nuclear piston mechanism consisting in pulling the nucleus via actomyosin contractility to push the cytoplasm that in turn push the membrane, leading to pseudopod formation. To occur, this lobopodia-based fibroblast migration needs nesprin 3, integrins, RhoA, ROCK and myosin II. Otherwise, lobopods are often accompanied with small lateral blebs forming along the side of the cell, probably due to the high intracellular pressure during lobopodia formation increasing the frequency of plasma membrane-cortex rupture.[18][7][19]
Reticulopodia
Reticulopodia (or reticulose pseudopods),[20] are complex formations in which individual pseudopods are merged and form irregular nets. The primary function of reticulopodia, also known as myxopodia, is food ingestion, with locomotion a secondary function. Reticulopods are typical of Foraminifera, Chlorarachnea, Gromia and Filoreta (Rhizaria).[4]
Axopodia
Axopodia (also known as actinopodia) are narrow pseudopodia containing complex arrays of microtubules enveloped by cytoplasm. Axopodia are mostly responsible for phagocytosis by rapidly retracting in response to physical contact. Principally, these pseudopodia are food collecting structures. They are observed in "Radiolaria" and "Heliozoa".[4]
References
- Etienne-Manneville S (2004). "Actin and Microtubules in Cell Motility: Which One is in Control?". Traffic. 5: 470–77. doi:10.1111/j.1600-0854.2004.00196.x.
- Tang DD (2017). "The roles and regulation of the actin cytoskeleton, intermediate filaments and microtubules in smooth muscle cell migration". Respiratory Research. 18: 54. doi:10.1186/s12931-017-0544-7. PMC 5385055. PMID 28390425.
- Patterson, David J. "Amoebae: Protists Which Move and Feed Using Pseudopodia". Tree of Life Web Project. Retrieved 2017-11-12.
- "Pseudopodia". Arcella.nl. Archived from the original on 2018-12-16. Retrieved 2018-12-16.
- Xue F; et al. (2010). "Contribution of Filopodia to Cell Migration: A Mechanical Link between Protrusion and Contraction". Journal of Cell Biology. 2010: 1–13. doi:10.1155/2010/507821. PMC 2910478. PMID 20671957.
- Machesky LM; et al. (2008). "Lamellipodia and filopodia in metastasis and invasion". FEBS Letters. 582 (14): 2102–11. doi:10.1016/j.febslet.2008.03.039.
- Petrie RJ; et al. (2012). "Nonpolarized signaling reveals two distinct modes of 3D cell migration". Journal of Cell Biology. 197 (3): 439. doi:10.1083/jcb.201201124. PMC 3341168. PMID 22547408.
- Bogitsh, Burton J.; Carter, Clint E.; Oeltmann, Thomas N. (2013). "General Characteristics of the Euprotista (Protozoa)". Human Parasitology. pp. 37–51. doi:10.1016/B978-0-12-415915-0.00003-0. ISBN 978-0-12-415915-0.
- "Pseudopodia". Encyclopedia.com. Retrieved 2018-12-16.
- Bosgraaf L & Van Haastert PJM (2009). "The Ordered Extension of Pseudopodia by Amoeboid Cells in the Absence of External Cues". PLOS ONE. 4 (4): 626–634. doi:10.1371/journal.pone.0005253. PMC 2668753. PMID 19384419.
- Van Haastert PJM & Devreotes PN (2004). "Chemotaxis: signalling the way forward". Nature Reviews Molecular Cell Biology. 5: e5253. doi:10.1038/nrm1435.
- Campbell EJ (2017). "A computational model of amoeboid cell swimming". Physics of Fluids. 29: 101902. doi:10.1063/1.4990543.
- Conti MA (2008). "Nonmuscle myosin II moves in new directions". Journal of Cell Science. 121: 11–18. doi:10.1242/jcs.007112. PMID 18096687.
- Schoenwolf, Gary (2009). Larsen's Human Embryology (4th ed.). Churchill Livingstone Elsevier.
- Bray, Dennis (2001). Cell Movements: From molecules to motility second edition.
- Danijela Vignjevic; et al. (2006). "Role of fascin in filopodial protrusion". Journal of Cell Biology. 174 (6): 863–875. doi:10.1083/jcb.200603013. PMC 2064340. PMID 16966425.
- "Pseudopodium | cytoplasm". Encyclopedia Britannica. Retrieved 2018-12-16.
- Chengappa P; et al. (2018). "Chapter Seven - Intracellular Pressure: A Driver of Cell Morphology and Movement". International Review of Cell and Molecular Biology. 337: 185–211. doi:10.1016/bs.ircmb.2017.12.005.
- Petrie RJ; et al. (2017). "Activating the nuclear piston mechanism of 3D migration in tumor cells". Journal of Cell Biology. 216 (1): 93. doi:10.1083/jcb.201605097.
- "Reticulopodia". eForams. Archived from the original on 2007-07-17. Retrieved 2005-12-30.
| Authority control |
|
|---|