Phalloidin
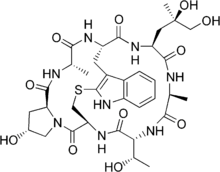
Phalloidin belongs to a class of toxins called phallotoxins, which are found in the death cap mushroom (Amanita phalloides). It is a rigid bicyclic heptapeptide that is lethal after a few days when injected into the bloodstream. The major symptom of phalloidin poisoning is acute hunger due to the destruction of liver cells. It functions by binding and stabilizing filamentous actin (F-actin) and effectively prevents the depolymerization of actin fibers. Due to its tight and selective binding to F-actin, derivatives of phalloidin containing fluorescent tags are used widely in microscopy to visualize F-actin in biomedical research. Though phallotoxins are highly toxic to liver cells, they add little to the toxicity of ingested death cap, as they are not absorbed through the gut.
 | |
| Identifiers | |
|---|---|
3D model (JSmol) |
|
| ChEBI | |
| ChemSpider | |
| ECHA InfoCard | 100.037.697 |
PubChem CID |
|
| UNII | |
CompTox Dashboard (EPA) |
|
| |
| |
| Properties | |
| C35H48N8O11S | |
| Molar mass | 788.87 g·mol−1 |
| Appearance | Needles |
| Melting point | 281 °C (538 °F; 554 K) (hyd) |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
Discovery and background
Phalloidin was one of the first cyclic peptides to be discovered. It was isolated from the death cap mushroom and crystallized by Feodor Lynen and Ulrich Wieland[1] in 1937.[2] Its structure is unusual in that it contains a cysteine-tryptophan linkage to form a bicyclic heptapeptide. This linkage had not been characterized before and makes the structure elucidation of phalloidin significantly more difficult. They determined the presence of the sulfur atom using UV spectroscopy and found that this ring structure had a slightly shifted wavelength. Raney nickel experiments confirmed the presence of sulfur in the tryptophan ring. The researchers found the desulfurized phalloidin was still circular, which demonstrated that the structure of phalloidin is normally bicyclic. Once linearized, the amino acid sequence of de-sulfurized phalloidin was elucidated through Edman degradation by Wieland and Schön in 1955.[3]
Due to its high affinity for actin, scientists discovered its potential use as a staining reagent for effective visualization of actin in microscopy. Derivatives conjugated with fluorophores are sold widely. Because of its ability to selectively bind filamentous actin (F-actin) and not actin monomers (G-actin), fluorescently labeled phalloidin is more effective than antibodies against actin.[4]
Synthesis
Biosynthesis
Phalloidin is a bicyclic heptapeptide containing an unusual cysteine-tryptophan linkage. The gene coding for synthesis of phalloidin is part of the MSDIN family in the Death Cap mushroom and codes for a 34 amino acid propeptide. A proline residue flanks the seven-residue region that will later become phalloidin. After translation, the peptide must be proteolyticly excised, cyclized, hydroxylated, Trp-Cys cross-linked to form tryptathionine, and epimerized to form a D-Thr. The order and exact biochemical mechanism for these steps is not yet fully understood. The current belief is that the necessary biosynthetic genes are clustered near the MSDIN genes.[5]
The first post-translational modification of the 34-mer is proteolytic cleavage via a prolyl oligopeptidase (POP) to remove the 10-amino acid "leader" peptide. The POP then cyclizes the heptapeptide Ala-Trp-Leu-Ala-Thr-Cys-Pro by transpeptidation between amino acid 1 (Ala) and amino acid 7 (Pro). It is believed that the formation of tryptathionine through Trp-Cys cross-linking occurs next.[5]
Chemical synthesis
Since phalloidin is exploited for its ability to bind and stabilize actin polymers but cells cannot readily uptake it, scientists have found phalloidin derivatives to be more useful in research. Essentially, it follows typical small peptide synthesis, using hydroxyl-proline. The major difficulty in synthesis is the formation of the tryptathionine bond (cysteine - tryptophan cross-linkage).
Below is the general synthetic mechanism carried out by Anderson et al. in 2005 for the solid phase synthesis of ala7-phalloidin, which differs at residue 7 from phalloidin as indicated below.[6] THPP stands for tetrahydropyranyl polystyrene linker, which is used to connect the molecule with the solid support during synthesis. Note that the synthesis below is simply a general scheme to show the order of bond formation to connect the starting materials. Ala7-phalloidin as well as many other similar variants of phalloidin are useful to increase cell uptake relative to phalloidin and to attach a fluorophore to aid in the visualization of F-actin in microscopy.
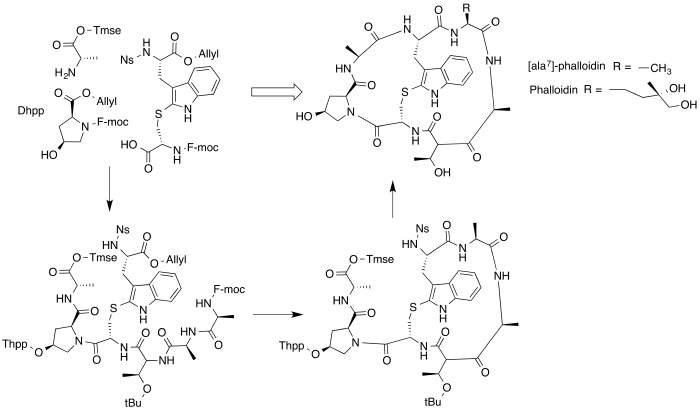
The first total synthesis of phalloidin was achieved through a combination of solid phase and solution phase synthesis (Baosheng Liu and Jianheng Zhang, United States Patent, US 8,569,452 B2). The physical and chemical properties of the synthetic phalloidin are the same as the naturally occurring phalloidin.
Mechanism of action
Phalloidin binds F-actin, preventing its depolymerization and poisoning the cell. Phalloidin binds specifically at the interface between F-actin subunits, locking adjacent subunits together. Phalloidin, a bicyclic heptapeptide, binds to actin filaments much more tightly than to actin monomers, leading to a decrease in the rate constant for the dissociation of actin subunits from filament ends, which essentially stabilizes actin filaments through the prevention of filament depolymerization.[7] Moreover, phalloidin is found to inhibit the ATP hydrolysis activity of F-actin.[8] Thus, phalloidin traps actin monomers in a conformation distinct from G-actin and it stabilizes the structure of F-actin by greatly reducing the rate constant for monomer dissociation, an event associated with the trapping of ADP.[8] Overall, phalloidin is found to react stoichiometrically with actin, strongly promote actin polymerization, and stabilize actin polymers.[9]
Phalloidin functions differently at various concentrations in cells. When introduced into the cytoplasm at low concentrations, phalloidin recruits the less polymerized forms of cytoplasmic actin as well as filamin into stable "islands" of aggregated actin polymers, yet it does not interfere with stress fibers, i.e. thick bundles of microfilaments.[9] Wehland et al. also notes that at higher concentrations, phalloidin induces cellular contraction.[9]
Symptoms
Soon after its discovery, scientists injected phalloidin into mice and discovered its LD50 is 2 mg/kg via IP injection. When exposed to the minimum lethal dose, it took several days for these mice to die. The only apparent side effect of phalloidin poisoning is extreme hunger. This is because phalloidin is only taken up by the liver via bile salt membrane transport proteins.[10] Once inside the liver, phalloidin binds F-actin, preventing its depolymerization. It takes time for this process to destroy the liver cells. The kidneys can also take up phalloidin, but not as effectively as the liver. Here, phalloidin causes nephrosis.[11]
Use as an imaging tool
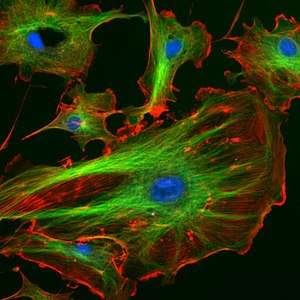
The properties of phalloidin make it a useful tool for investigating the distribution of F-actin in cells by labeling phalloidin with fluorescent analogs and using them to stain actin filaments for light microscopy. Fluorescent derivatives of phalloidin have turned out to be enormously useful in localizing actin filaments in living or fixed cells as well as for visualizing individual actin filaments in vitro.[7] A high-resolution technique was developed to detect F-actin at the light and electron microscopic levels by using phalloidin conjugated to the fluorophore eosin which acts as the fluorescent tag.[12] In this method known as fluorescence photo-oxidation, fluorescent molecules can be utilized to drive the oxidation of diaminobenzidine (DAB) to create a reaction product that can be rendered electron dense and detectable by electron microscopy.[12] The amount of fluorescence visualized can be used as a quantitative measure of the amount of filamentous actin there is in cells if saturating quantities of fluorescent phalloidin are used.[7] Consequently, immunofluorescence microscopy along with microinjection of phalloidin can be used to evaluate the direct and indirect functions of cytoplasmic actin in its different stages of polymer formation.[9] Therefore, fluorescent phalloidin can be used as an important tool in the study of actin networks at high resolution.
Uses and limitations
Phalloidin is much smaller than an antibody that would typically be used to label cellular proteins for fluorescent microscopy which allows for much denser labeling of filamentous actin and much more detailed images can be acquired particularly at higher resolutions.
Unmodified phalloidins do not permeate cell membranes, making them less effective in experiments with living cells. Derivatives of phalloidin with greatly increased cell permeability have been synthesized.
Cells treated with phalloidins exhibit a number of toxic effects and frequently die.[7] Furthermore, it is important to note that phalloidin-treated cells will have greater levels of actin associated with their plasma membranes, and the microinjection of phalloidin into living cells will change actin distribution as well as cell motility.[7]
See also
References
- G. Semenza, E.C. Slater, R. Jaenicke. Selected Topics in the History of Biochemistry. Personal Recollections - Google books
- Lynen F, Wieland U (18 November 1937). "Uber die Giftstoffe des Knollenblätterpilzes. IV". Justus Liebigs Annalen der Chemie (in German). 533 (1): 93–117. doi:10.1002/jlac.19385330105.
- Wieland T, Schon W (16 January 1955). "Über die Giftstoffe des grünen Knollenblätterpilzes X. Mitteilung. Die Konstitution des Phalloidins". Justus Liebigs Annalen der Chemie. 593 (2): 157–178. doi:10.1002/jlac.19555930204.
- Immunohistochemistry: Basics and Methods. Springer Science & Business Media. 2010. pp. 92–3. ISBN 978-3-642-04609-4.
- Walton JD; Hallen-Adams He; Luo H (4 August 2010). "Ribosomal biosynthesis of the cyclic peptide toxins of Amanita mushrooms". Peptide Science. 94 (5): 659–664. doi:10.1002/bip.21416. PMC 4001729. PMID 20564017.
- Anderson MO, Shelat AA, Kiplan Guy R (16 April 2005). "A solid-phase approach to the phallotoxins: total synthesis of [ala7]-phalloidin". J. Org. Chem. 70 (12): 4578–84. doi:10.1021/jo0503153. PMID 15932292.
- Cooper JA (October 1987). "Effects of cytochalasin and phalloidin on actin". J. Cell Biol. 105 (4): 1473–8. doi:10.1083/jcb.105.4.1473. PMC 2114638. PMID 3312229.
- Barden JA, Miki M, Hambly BD, Dos Remedios CG (February 1987). "Localization of the phalloidin and nucleotide-binding sites on actin". Eur. J. Biochem. 162 (3): 583–8. doi:10.1111/j.1432-1033.1987.tb10679.x. PMID 3830158.
- Wehland J, Osborn M, Weber K (December 1977). "Phalloidin-induced actin polymerization in the cytoplasm of cultured cells interferes with cell locomotion and growth". Proc. Natl. Acad. Sci. U.S.A. 74 (12): 5613–7. doi:10.1073/pnas.74.12.5613. PMC 431831. PMID 341163.
- Wieland T (1963). "Chemical and toxicological studies with cyclopeptides of Amanita phalloides". Pure and Applied Chemistry. 3 (6): 339–350. doi:10.1351/pac196306030339.
- Schröder, Eberhard; Lübke, Klaus (2014). The Peptides, Volume II: Synthesis, Occurrence, and Action of Biologically Active Polypeptides. Elsevier. p. 475. ISBN 978-1-4832-5986-4. Focuses on the synthesis of biologically active polypeptides and analogues.
- Capani F, Deerinck TJ, Ellisman MH, Bushong E, Bobik M, Martone ME (1 November 2001). "Phalloidin-eosin followed by photo-oxidation: a novel method for localizing F-actin at the light and electron microscopic levels". J. Histochem. Cytochem. 49 (11): 1351–61. doi:10.1177/002215540104901103. PMID 11668188. Archived from the original on 16 April 2005.
Poisonous Amanita mushrooms | |||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Subgenus Amanita |
|  | |||||||||||||||||||||||||||||||||||
| Subgenus Amanitina |
| ||||||||||||||||||||||||||||||||||||