Lyme disease microbiology
Lyme disease, or borreliosis, is caused by spirochetal bacteria from the genus Borrelia,[1] which has 52 known species. Three main species (Borrelia garinii, Borrelia afzelii, and Borrelia burgdorferi s.s.) are the main causative agents of the disease in humans,[2] while a number of others have been implicated as possibly pathogenic.[3][4] Borrelia species in the species complex known to cause Lyme disease are collectively called Borrelia burgdorferi sensu lato (s.l.) not to be confused with the single species in that complex Borrelia burgdorferi sensu stricto which is responsible for all cases of Lyme disease in North America.[5]
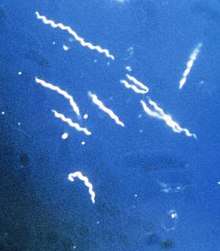
Borrelia are microaerophilic and slow-growing—the primary reason for the long delays when diagnosing Lyme disease—and have been found to have greater strain diversity than previously estimated.[6] The strains differ in clinical symptoms and/or presentation as well as geographic distribution.[7]
Except for Borrelia recurrentis (which causes louse-borne relapsing fever and is transmitted by the human body louse), all known species are believed to be transmitted by ticks.[8]
Species and strains
Until recently, only three genospecies were thought to cause Lyme disease (borreliosis): B. burgdorferi s.s. (the predominant species in North America, but also present in Europe); B. afzelii; and B. garinii (both predominant in Eurasia).
Thirteen distinct genomic classifications of Lyme disease bacteria have been identified worldwide. These include but are not limited to B. burgdorferi s.s., B. afzelii, B. garinii, B. valaisana, B. lusitaniae, B. andersoni, 25015, DN127, CA55, 25015, HK501, B. miyamotoi, and B. japonica.[9] Many of these genomic groups are country or continent specific. For example, without migration, B. japonica is only prevalent in the eastern hemisphere.[9]
The genomic variations have direct implications on the clinical symptoms of tick-borne Lyme disease. For example, B. burgdorferi s.s.’s tick-borne Lyme disease may manifest with arthritis-like symptoms.[9] In contrast, B. garinii’s tick-borne Lyme disease may cause an infection of the central nervous system.[9]
Emerging genospecies
- B. valaisiana was identified as a genomic species from Strain VS116, and named B. valaisiana in 1997.[10] It was later detected by polymerase chain reaction (PCR) in human cerebral spinal fluid (CSF) in Greece.[11] B. valaisiana has been isolated throughout Europe, as well as east Asia.[12]
Newly discovered genospecies have also been found to cause disease in humans:
- B. lusitaniae[13] in Europe (especially Portugal), North Africa and Asia.
- B. bissettii[14][15] in the United States and Europe.
- B. spielmanii[16][17] in Europe.
Additional B. burgdorferi sensu lato genospecies suspected of causing illness, but not confirmed by culture, include B. japonica, B. tanukii and B. turdae (Japan); B. sinica (China); and B. andersonii (U.S.). Some of these species are carried by ticks not currently recognized as carriers of Lyme disease.
The B. miyamotoi spirochete, related to the relapsing fever group of spirochetes, is also suspected of causing illness in Japan. Spirochetes similar to B. miyamotoi have recently been found in both Ixodes ricinus ticks in Sweden and I. scapularis ticks in the U.S.[18][19][20]
Epidemiology
Lyme disease is most endemic in Northern Hemisphere temperate regions,[21][22] but sporadic cases have been described in other areas of the world.
The number of reported cases of the borreliosis have been increasing, as are endemic regions in North America. Of cases reported to the United States Centers for Disease Control and Prevention (CDC), the rate of Lyme disease infection is 7.9 cases for every 100,000 persons. In the 10 states where Lyme disease is most common, the average was 31.6 cases per 100,000 persons for 2005.[23] Although Lyme disease has now been reported in 49 of 50 states in the U.S (all but Hawaii), about 99% of all reported cases are confined to just five geographic areas (New England, Mid-Atlantic, East-North Central, South Atlantic, and West North-Central).[24]
In Europe, cases of B. burgdorferi s.l.-infected ticks are found predominantly in Norway, the Netherlands, Germany, France, Italy, Slovenia, and Poland, but have been isolated in almost every country on the continent. Lyme disease statistics for Europe can be found at Eurosurveillance website.
Borrelia burgdorferi s.l.-infested ticks are being found more frequently in Japan, as well as in northwest China and far eastern Russia.[25][26] Borrelia has been isolated in Mongolia as well.[27]
In South America, tick-borne disease recognition and occurrence is rising. Ticks carrying B. burgdorferi s.l., as well as canine and human tick-borne diseases, have been reported widely in Brazil, but the subspecies of Borrelia has not yet been defined.[28] The first reported case of Lyme disease in Brazil was made in 1993 in Sao Paulo.[29] B. burgdorferi sensu stricto antigens in patients have been identified in Colombia and in Bolivia. B. burgorferi has been reported in Bay Islands of Honduras.
In Northern Africa, B. burgdorferi s.s. has been identified in Morocco, Algeria, Egypt, and Tunisia.[30][31][32]
In western and sub-Saharan Africa, tick-borne relapsing fever has been recognized for over a century, since it was first isolated by the British physicians Joseph Everett Dutton and John Lancelot Todd in 1905. Borrelia in the manifestation of Lyme disease in this region is presently unknown, but evidence indicates the disease may occur in humans in sub-Saharan Africa. The abundance of hosts and tick vectors would favor the establishment of the infection in Africa.[33] In East Africa two cases of Lyme disease have been reported in Kenya.[34]
In Australia, no definitive evidence exists for the existence of B. burgdorferi or for any other tick-borne spirochete that may be responsible for a local syndrome being reported as Lyme disease.[35] Cases of neuroborreliosis have been documented in Australia, but are often ascribed to travel to other continents. The existence of Lyme disease in Australia is controversial.[36]
Lifecycle
The lifecycle of B. burgdorferi is complex, requiring ticks, and species that are competent reservoirs, often small rodents. Mice are the primary reservoir for the bacteria.
Hard ticks have a variety of life histories with respect to optimizing their chance of contact with an appropriate host to ensure survival. The life stages of soft ticks are not readily distinguishable. The first stage to hatch from the egg, a six-legged larva, takes a blood meal from a host, and molts to the first nymphal stage. Unlike hard ticks, many soft ticks go through multiple nymphal stages, gradually increasing in size until the final molt to the adult stage.
The lifecycle of the black-legged tick, commonly called the deer tick (Ixodes scapularis) comprises three growth stages: the larva, nymph and adult.
Whereas B. burgdorferi is most associated with deer ticks and the white-footed mouse,[37] B. afzelli is most frequently detected in rodent-feeding vector ticks, and B. garinii and B. valaisiana appear to be associated with birds. Both rodents and birds are competent reservoir hosts for B. burgdorferi sensu stricto. The resistance of a genospecies of Lyme disease spirochetes to the bacteriolytic activities of the alternative immune complement system of various host species may determine its reservoir host association.
Genomic characteristics
The genome of B. burgdorferi (B31 strain) was the third microbial genome ever to be sequenced, following the sequencing of both H. influenzae and M. genitalium in 1995, and its chromosome contains 910,725 base pairs and 853 genes.[38] One of the most striking features of B. burgdorferi as compared with other bacteria is its unusual genome, which is far more complex than that of its spirochetal cousin Treponema pallidum, the agent of syphilis.[39] In addition to a linear chromosome, the genome of B. burgdorferi strain B31 includes 21 plasmids (12 linear and 9 circular) – by far the largest number of plasmids found in any known bacterium.[40] Genetic exchange, including plasmid transfers, contributes to the pathogenicity of the organism.[41] Long-term culture of B. burgdorferi results in a loss of some plasmids and changes in expressed protein profiles. Associated with the loss of plasmids is a loss in the ability of the organism to infect laboratory animals, suggesting the plasmids encode key genes involved in virulence.
Chemical analysis of the external membrane of B. burgdorferi revealed the presence of 46% proteins, 51% lipids and 3% carbohydrates.[42]
Structure and growth
B. burgdorferi is a highly specialized, motile, two-membrane, flat-waved spirochete, ranging from about 9 to 32 μm in length.[43] Because of its double-membrane envelope, it is often mistakenly described as Gram negative,[44] though it stains weakly in Gram stain. The bacterial membranes in at least the B31, NL303 and N40 strains of B. burgdorferi do not contain lipopolysaccharide, which is extremely atypical for Gram negative bacteria; instead, the membranes contain glycolipids.[45] However, the membranes in the B31 strain have been found to contain a lipopolysaccharide-like component.[46] B. burgdorferi is a microaerophilic organism, requiring little oxygen to survive. Unlike most bacteria, B. burgdorferi does not use iron, hence avoiding the difficulty of acquiring iron during infection.[47] It lives primarily as an extracellular pathogen.
Like other spirochetes, such as Treponema pallidum (the agent of syphilis), B. burgdorferi has an axial filament composed of flagella that run lengthways between its cell wall and outer membrane. This structure allows the spirochete to move efficiently in corkscrew fashion through viscous media, such as connective tissue.
B. burgdorferi is very slow growing, with a doubling time of 12–18 hours[48] (in contrast to pathogens such as Streptococcus and Staphylococcus, which have a doubling time of 20–30 minutes).
Outer surface proteins
The outer membrane of B. burgdorferi is composed of various unique outer surface proteins (Osp) that have been characterized (OspA through OspF). The Osp proteins are lipoproteins anchored by N-terminally attached fatty acid molecules to the membrane.[49] They are presumed to play a role in virulence, transmission, or survival in the tick.
OspA, OspB, and OspD are expressed by B. burgdorferi residing in the gut of unfed ticks, suggesting they promote the persistence of the spirochete in ticks between blood meals.[50][51] During transmission to the mammalian host, when the nymphal tick begins to feed and the spirochetes in the midgut begin to multiply rapidly, most spirochetes cease expressing OspA on their surfaces. Simultaneous with the disappearance of OspA, the spirochete population in the midgut begins to express an OspC and migrates to the salivary gland. Upregulation of OspC begins during the first day of feeding and peaks 48 hours after attachment.[52]
The OspA and OspB genes encode the major outer membrane proteins of B. burgdorferi. The two Osp proteins show a high degree of sequence similarity, indicating a recent duplication event.[53] Virtually all spirochetes in the midgut of an unfed nymph tick express OspA. OspA promotes the attachment of B. burgdorferi to the tick protein TROSPA, present on tick gut epithelial cells.[54] OspB also has an essential role in the adherence of B. burgdorferi to the tick gut.[55] Although OspD has been shown to bind to tick gut extracts in vitro, as well as OspA and OspB, it is not essential for the attachment and colonization of the tick gut, and it is not required for human infections.[51]
OspC is a strong antigen; detection of its presence by the host organism stimulates an immune response. While each individual bacterial cell contains just one copy of the ospC gene, the gene sequence of ospC among different strains within each of the three major Lyme disease species is highly variable.[56] OspC plays an essential role during the early stage of mammalian infection.[57] In infected ticks feeding on a mammalian host, OspC may also be necessary to allow B. burgdorferi to invade and attach to the salivary gland after leaving the gut, although not all studies agree on such a role for the protein.[58][59] OspC attaches to the tick salivary protein Salp15, which protects the spirochete from complement and impairs the function of dendritic cells.[60][61][62]
OspE and OspF were initially identified in B. burgdorferi strain N40.[63] The ospE and ospF genes are structurally arranged in tandem as one transcriptional unit under the control of a common promoter.[63] Individual strains of B. burgdorferi carry multiple related copies of the ospEF locus, which are now collectively referred to as Erp (Osp'E'/F-like 'r'elated 'p'rotein) genes. In B. burgdoreri strains B31 and 297, most of the Erp loci occupy the same position on the multiple copies of the cp32 plasmid present in these strains.[64] Each locus consists of one or two Erp genes. When two genes are present, they are transcribed as one operon, although in some cases, an internal promoter in the first gene may also transcribe the second gene.[65] The presence of multiple Erp proteins was proposed to be important in allowing B. burgdorferi to evade killing by the alternative complement pathway of a broad range of potential animal hosts, as individual Erp proteins exhibited different binding patterns to the complement regulator factor H from different animals.[66] However, the presence of factor H was recently demonstrated to not be necessary to enable B. burgdorferi to infect mice, suggesting the Erp proteins have an additional function.[67]
Mechanisms of persistence
B. burgdorferi is susceptible to a number of antibiotics in humans. However, untreated B. burgdorferi may persist in humans for months or years. In North America and Europe, Lyme arthritis may persist, while Europe also includes a persistent skin condition called acrodermatitis chronica atrophicans.[68]
- Antigenic variation and gene expression
Like the Borrelia that causes relapsing fever, B. burgdorferi has the ability to vary its surface proteins in response to immune attack.[69][70] This ability is related to the genomic complexity of B. burgdorferi, and is another way B. burgdorferi evades the immune system to establish a chronic infection.[71]
References
- Samuels DS, Radolf JD, eds. (2010). Borrelia: Molecular Biology, Host Interaction and Pathogenesis. Caister Academic Press. ISBN 978-1-904455-58-5.
- Stanek, Gerold; Wormser, Gary P; Gray, Jeremy; Strle, Franc (7 September 2011). "Lyme borreliosis". The Lancet. 379 (9814): 461–73. doi:10.1016/S0140-6736(11)60103-7. PMID 21903253.
- Cutler SJ, Ruzic-Sabljic E, Potkonjak A (2016). "Emerging borreliae - Expanding beyond Lyme borreliosis" (PDF). Molecular and Cellular Probes. 31: 22–27. doi:10.1016/j.mcp.2016.08.003. PMID 27523487.
- Pritt, Bobbi S (5 February 2016). "Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: a descriptive study". The Lancet Infectious Diseases. 16 (5): 556–564. doi:10.1016/S1473-3099(15)00464-8. PMC 4975683. PMID 26856777.
- Tilly, Kit; Rosa, Patricia A.; Stewart, Philip E. (2008). "Biology of Infection with Borrelia burgdorferi". Infectious Disease Clinics of North America. 22 (2): 217–234. doi:10.1016/j.idc.2007.12.013. PMC 2440571. PMID 18452798.
- Bunikis J, Garpmo U, Tsao J, Berglund J, Fish D, Barbour AG (2004). "Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe" (PDF). Microbiology. 150 (Pt 6): 1741–55. doi:10.1099/mic.0.26944-0. PMID 15184561.
- Ryan KJ, Ray CG, eds. (2004). Sherris Medical Microbiology (4th ed.). McGraw Hill. ISBN 978-0-8385-8529-0.
- Felsenfeld O (1971). Borrelia: Strains, Vectors, Human and Animal Borreliosis. St. Louis: Warren H. Green, Inc.
- Habálek, Z.; Halouzka, J. (1997-12-01). "Distribution of Borrelia burgdorferi sensu lato genomic groups in Europe, a review". European Journal of Epidemiology. 13 (8): 951–957. doi:10.1023/A:1007426304900. ISSN 0393-2990. PMID 9476827.
- Wang G, van Dam AP, Le Fleche A, et al. (1997). "Genetic and phenotypic analysis of Borrelia valaisiana sp. nov. (Borrelia genomic groups VS116 and M19)". Int. J. Syst. Bacteriol. 47 (4): 926–932. doi:10.1099/00207713-47-4-926. PMID 9336888.
- Diza E, Papa A, Vezyri E, Tsounis S, Milonas I, Antoniadis A (2004). "Borrelia valaisiana in cerebrospinal fluid". Emerging Infect. Dis. 10 (9): 1692–3. doi:10.3201/eid1009.030439. PMC 3320289. PMID 15503409.
- Masuzawa T (2004). "Terrestrial distribution of the Lyme borreliosis agent Borrelia burgdorferi sensu lato in East Asia". Jpn. J. Infect. Dis. 57 (6): 229–235. PMID 15623946.
- Collares-Pereira M, Couceiro S, Franca I, Kurtenbach K, Schafer SM, Vitorino L, Goncalves L, Baptista S, Vieira ML, Cunha C (2004). "First isolation of Borrelia lusitaniae from a human patient" (PDF). J Clin Microbiol. 42 (3): 1316–8. doi:10.1128/JCM.42.3.1316-1318.2004. PMC 356816. PMID 15004107.
- Postic D, Ras NM, Lane RS, Hendson M, Baranton G (1998). "Expanded diversity among Californian Borrelia isolates and description of Borrelia bissettii sp. nov. (formerly Borrelia group DN127)" (PDF). J Clin Microbiol. 36 (12): 3497–3504. PMC 105228. PMID 9817861.
- Maraspin V, Cimperman J, Lotric-Furlan S, Ruzic-Sabljic E, Jurca T, Picken RN, Strle F (2002). "Solitary borrelial lymphocytoma in adult patients". Wien Klin Wochenschr. 114 (13–14): 515–523. PMID 12422593.
- Richter D, Postic D, Sertour N, Livey I, Matuschka FR, Baranton G (2006). "Delineation of Borrelia burgdorferi sensu lato species by multilocus sequence analysis and confirmation of the delineation of Borrelia spielmanii sp. nov". Int J Syst Evol Microbiol. 56 (Pt 4): 873–881. doi:10.1099/ijs.0.64050-0. PMID 16585709.
- Foldvari G, Farkas R, Lakos A (2005). "Borrelia spielmanii erythema migrans, Hungary". Emerg Infect Dis. 11 (11): 1794–5. doi:10.3201/eid1111.050542. PMC 3367353. PMID 16422006.
- Scoles GA, Papero M, Beati L, Fish D (2001). "A relapsing fever group spirochete transmitted by Ixodes scapularis ticks". Vector-Borne and Zoonotic Diseases. 1 (1): 21–34. doi:10.1089/153036601750137624. PMID 12653133.
- Bunikis J, Tsao J, Garpmo U, Berglund J, Fish D, Barbour AG (2004). "Typing of Borrelia relapsing fever group strains". Emerg Infect Dis. 10 (9): 1661–4. doi:10.3201/eid1009.040236. PMC 3320305. PMID 15498172.
- McNeil, Donald (19 September 2011). "New Tick-Borne Disease Is Discovered". The New York Times. pp. D6. Retrieved 20 September 2011.
- Grubhoffer L, Golovchenko M, Vancova M, Zacharovova-Slavickova K, Rudenko N, Oliver JH Jr (November 2005). "Lyme borreliosis: insights into tick-/host-borrelia relations". Folia Parasitol (Praha). 52 (4 (Review)): 279–294. doi:10.14411/fp.2005.039. PMID 16405291.
- Higgins R (August 2004). "Emerging or re-emerging bacterial zoonotic diseases: bartonellosis, leptospirosis, Lyme borreliosis, plague". Rev. Sci. Tech. 23 (2): 569–581. doi:10.20506/rst.23.2.1503. PMID 15702720.
- "DVBID: Disease Upward Climb – CDC Lyme Disease". 2006-10-02. Retrieved 2007-08-23.
- "Lyme Disease Statistics". Centers for Disease Control and Prevention (CDC). 2007-04-02. Retrieved 2007-08-23.
- Li M, Masuzawa T, Takada N, Ishiguro F, Fujita H, Iwaki A, Wang H, Wang J, Kawabata M, Yanagihara Y (July 1998). "Lyme disease Borrelia species in northeastern China resemble those isolated from far eastern Russia and Japan". Appl Environ Microbiol. 64 (7): 2705–9. doi:10.1128/AEM.64.7.2705-2709.1998. PMC 106449. PMID 9647853.
- Masuzawa T (December 2004). "Terrestrial distribution of the Lyme borreliosis agent Borrelia burgdorferi sensu lato in East Asia". Jpn J Infect Dis. 57 (6): 229–235. PMID 15623946.
- Walder G, Lkhamsuren E, Shagdar A, Bataa J, Batmunkh T, Orth D, Heinz FX, Danichova GA, Khasnatinov MA, Wurzner R, Dierich MP (May 2006). "Serological evidence for tick-borne encephalitis, borreliosis, and human granulocytic anaplasmosis in Mongolia". Int J Med Microbiol. 296 (Suppl 40): 69–75. doi:10.1016/j.ijmm.2006.01.031. PMID 16524782.
- Mantovani E, Costa IP, Gauditano G, Bonoldi VL, Higuchi ML, Yoshinari NH (April 2007). "Description of Lyme disease-like syndrome in Brazil: is it a new tick-borne disease or Lyme disease variation?". Braz J Med Biol Res. 40 (4): 443–456. doi:10.1590/S0100-879X2006005000082. PMID 17401487.
- Yoshinari NH, Oyafuso LK, Monteiro FG, de Barros PJ, da Cruz FC, Ferreira LG, Bonasser F, Baggio D, Cossermelli W (Jul–Aug 1993). "Lyme disease. Report of a case observed in Brazil". Rev Hosp Clin Fac Med Sao Paulo. 48 (4): 170–4. PMID 8284588.
- Bouattour A, Ghorbel A, Chabchoub A, Postic D (2004). "Lyme borreliosis situation in North Africa". Arch Inst Pasteur Tunis. 81 (1–4): 13–20. PMID 16929760.
- Dsouli N, Younsi-Kabachii H, Postic D, Nouira S, Gern L, Bouattour A (July 2006). "Reservoir role of lizard Psammodromus algirus in transmission cycle of Borrelia burgdorferi sensu lato (Spirochaetaceae) in Tunisia" (PDF). Journal of Medical Entomology. 43 (4): 737–742. doi:10.1603/0022-2585(2006)43[737:RROLPA]2.0.CO;2. ISSN 0022-2585. PMID 16892633.
- Helmy N (August 2000). "Seasonal abundance of Ornithodoros (O.) savignyi and prevalence of infection with Borrelia spirochetes in Egypt". J Egypt Soc Parasitol. 30 (2): 607–619. PMID 10946521.
- Fivaz BH, Petney TN (September 1989). "Lyme disease — a new disease in southern Africa?". J S Afr Vet Assoc. 60 (3): 155–8. PMID 2699499.
- Jowi JO, Gathua SN (May 2005). "Lyme disease: report of two cases". East Afr Med J. 82 (5): 267–9. doi:10.4314/eamj.v82i5.9318. PMID 16119758.
- Piesman J, Stone BF (February 1991). "Vector competence of the Australian paralysis tick, Ixodes holocyclus, for the Lyme disease spirochete Borrelia burgdorferi". Int J Parasitol. 21 (1): 109–111. doi:10.1016/0020-7519(91)90127-S. PMID 2040556.
- "Lyme: a four letter word". ABC Radio National Background Briefing. Australian Broadcasting Corporation. 12 May 2013. Retrieved 12 May 2013.
- Wallis RC, Brown SE, Kloter KO, Main AJ Jr (October 1978). "Erythema chronicum migrans and Lyme arthritis: field study of ticks". Am J Epidemiol. 108 (4): 322–7. doi:10.1093/oxfordjournals.aje.a112626. PMID 727201.
- Fraser, Claire M.; Casjens, S; Huang, WM; Sutton, GG; Clayton, R; Lathigra, R; White, O; Ketchum, KA; et al. (1997). "Genomic sequence of a Lyme disease spirochaete, Borrelia burgdorferi". Nature. 390 (6660): 580–6. doi:10.1038/37551. PMID 9403685.
- Porcella SF, Schwan TG (2001). "Borrelia burgdorferi and Treponema pallidum: a comparison of functional genomics, environmental adaptations, and pathogenic mechanisms". J Clin Invest. 107 (6): 651–6. doi:10.1172/JCI12484. PMC 208952. PMID 11254661.
- Casjens S, Palmer N, van Vugt R, Huang WM, Stevenson B, Rosa P, Lathigra R, Sutton G, Peterson J, Dodson RJ, Haft D, Hickey E, Gwinn M, White O, Fraser CM (2000). "A bacterial genome in flux: the twelve linear and nine circular extrachromosomal DNAs in an infectious isolate of the Lyme disease spirochete Borrelia burgdorferi". Mol Microbiol. 35 (3): 490–516. doi:10.1046/j.1365-2958.2000.01698.x. PMID 10672174.
- Qiu WG, Schutzer SE, Bruno JF, Attie O, Xu Y, Dunn JJ, Fraser CM, Casjens SR, Luft BJ (2004). "Genetic exchange and plasmid transfers in Borrelia burgdorferi sensu stricto revealed by three-way genome comparisons and multilocus sequence typing" (PDF). Proc Natl Acad Sci USA. 101 (39): 14150–5. doi:10.1073/pnas.0402745101. PMC 521097. PMID 15375210.
- Schwarzová K (June 1993). "Lyme borreliosis: review of present knowledge". Cesk Epidemiol Mikrobiol Imunol. 42 (2): 87–92. PMID 8348630.
- Goldstein SF, Charon NW, Kreiling JA (1994). "Borrelia burgdorferi swims with a planar waveform similar to that of eukaryotic flagella". Proc. Natl. Acad. Sci. U.S.A. 91 (8): 3433–7. doi:10.1073/pnas.91.8.3433. PMC 43591. PMID 8159765.
- Samuels DS, Radolf JD, eds. (2010). "Ch. 6: Structure, Function and Biogenesis of the Borrelia Cell Envelope". Borrelia: Molecular Biology, Host Interaction and Pathogenesis. Caister Academic Press. ISBN 978-1-904455-58-5.
- Ben-Menachem G, Kubler-Kielb J, Coxon B, Yergey A, Schneerson R (2003). "A newly discovered cholesteryl galactoside from Borrelia burgdorferi". Proc. Natl. Acad. Sci. U.S.A. 100 (13): 7913–8. doi:10.1073/pnas.1232451100. PMC 164687. PMID 12799465.
- Schwarzová K, Čižnár I (2004). "Immunochemical analysis of lipopolysaccharide-like component extracted from Borrelia burgdorferi sensu lato" (PDF). Folia Microbiol. 49 (5): 625–9. doi:10.1007/BF02931545. PMID 15702557. Archived from the original (PDF) on 2011-07-21. Retrieved 2007-10-26.
- Posey JE, Gherardini FC (2000). "Lack of a role for iron in the Lyme disease pathogen". Science. 288 (5471): 1651–3. doi:10.1126/science.288.5471.1651. PMID 10834845.
- Kelly, RT (1984). Krieg NR, Holt JG (eds.). Genus IV. Borrelia Swellengrebel 1907, 582AL. 1. Williams & Wilkins: Baltimore. pp. 57–62.
- Haake DA (2000). "Spirochaetal lipoproteins and pathogenesis". Microbiology. 146 (7): 1491–1504. doi:10.1099/00221287-146-7-1491. PMC 2664406. PMID 10878114.
- Schwan TG, Piesman J, Golde WT, Dolan MC, Rosa PA (1995). "Induction of an outer surface protein on Borrelia burgdorferi during tick feeding". Proc. Natl. Acad. Sci. U.S.A. 92 (7): 2909–13. doi:10.1073/pnas.92.7.2909. PMC 42328. PMID 7708747.
- Li X, Neelakanta G, Liu X, Beck DS, Kantor FS, Fish D, Anderson JF, Fikrig E (2007). "Role of outer surface protein D in the Borrelia burgdorferi life cycle". Infect. Immun. 75 (9): 4237–44. doi:10.1128/IAI.00632-07. PMC 1951184. PMID 17620358.
- Schwan TG, Piesman J (2000). "Temporal changes in outer surface proteins A and C of the Lyme disease-associated spirochete, Borrelia burgdorferi, during the chain of infection in ticks and mice". J Clin Microbiol. 38 (1): 382–8. PMC 88728. PMID 10618120.
- Bergström S, Bundoc VG, Barbour AG (1989). "Molecular analysis of linear plasmid-encoded major surface proteins, OspA and OspB, of the Lyme disease spirochaete Borrelia burgdorferi". Mol. Microbiol. 3 (4): 479–486. doi:10.1111/j.1365-2958.1989.tb00194.x. PMID 2761388.
- Pal U, Li X, Wang T, Montgomery RR, Ramamoorthi N, Desilva AM, Bao F, Yang X, Pypaert M, Pradhan D, Kantor FS, Telford S, Anderson JF, Fikrig E (2004). "TROSPA, an Ixodes scapularis receptor for Borrelia burgdorferi". Cell. 119 (4): 457–468. doi:10.1016/j.cell.2004.10.027. PMID 15537536.
- Neelakanta G, Li X, Pal U, Liu X, Beck DS, DePonte K, Fish D, Kantor FS, Fikrig E (2007). "Outer surface protein B is critical for Borrelia burgdorferi adherence and survival within Ixodes ticks". PLoS Pathog. 3 (3): e33. doi:10.1371/journal.ppat.0030033. PMC 1817655. PMID 17352535.
- Baranton G, Seinost G, Theodore G, Postic D, Dykhuizen D (March 2001). "Distinct levels of genetic diversity of Borrelia burgdorferi are associated with different aspects of pathogenicity". Res. Microbiol. 152 (2): 149–56. doi:10.1016/S0923-2508(01)01186-X. PMID 11316368.
- Tilly K, Krum JG, Bestor A, Jewett MW, Grimm D, Bueschel D, Byram R, Dorward D, Vanraden MJ, Stewart P, Rosa P (June 2006). "Borrelia burgdorferi OspC protein required exclusively in a crucial early stage of mammalian infection". Infect. Immun. 74 (6): 3554–64. doi:10.1128/IAI.01950-05. PMC 1479285. PMID 16714588.
- Pal U, Yang X, Chen M, Bockenstedt LK, Anderson JF, Flavell RA, Norgard MV, Fikrig E (January 2004). "OspC facilitates Borrelia burgdorferi invasion of Ixodes scapularis salivary glands". J. Clin. Invest. 113 (2): 220–30. doi:10.1172/JCI19894. PMC 311436. PMID 14722614.
- Grimm D, Tilly K, Byram R, Stewart PE, Krum JG, Bueschel DM, Schwan TG, Policastro PF, Elias AF, Rosa PA (March 2004). "Outer-surface protein C of the Lyme disease spirochete: a protein induced in ticks for infection of mammals". Proc. Natl. Acad. Sci. U.S.A. 101 (9): 3142–7. doi:10.1073/pnas.0306845101. PMC 365757. PMID 14970347.
- Ramamoorthi N, Narasimhan S, Pal U, Bao F, Yang XF, Fish D, Anguita J, Norgard MV, Kantor FS, Anderson JF, Koski RA, Fikrig E (July 2005). "The Lyme disease agent exploits a tick protein to infect the mammalian host". Nature. 436 (7050): 573–7. doi:10.1038/nature03812. PMC 4306560. PMID 16049492.
- Schuijt TJ, Hovius JW, van Burgel ND, Ramamoorthi N, Fikrig E, van Dam AP (July 2008). "The tick salivary protein Salp15 inhibits the killing of serum-sensitive Borrelia burgdorferi sensu lato isolates". Infect. Immun. 76 (7): 2888–94. doi:10.1128/IAI.00232-08. PMC 2446733. PMID 18426890.
- Hovius JW, de Jong MA, den Dunnen J, Litjens M, Fikrig E, van der Poll T, Gringhuis SI, Geijtenbeek TB (February 2008). "Salp15 binding to DC-SIGN inhibits cytokine expression by impairing both nucleosome remodeling and mRNA stabilization". PLoS Pathog. 4 (2): e31. doi:10.1371/journal.ppat.0040031. PMC 2242833. PMID 18282094.
- Lam TT, Nguyen TP, Montgomery RR, Kantor FS, Fikrig E, Flavell RA (1994). "Outer surface proteins E and F of Borrelia burgdorferi, the agent of Lyme disease". Infect. Immun. 62 (1): 290–8. PMC 186099. PMID 8262642.
- Stevenson B, Zückert WR, Akins DR (2000). "Repetition, conservation, and variation: the multiple cp32 plasmids of Borrelia species". J. Mol. Microbiol. Biotechnol. 2 (4): 411–422. PMID 11075913.
- Stevenson B, Bono JL, Schwan TG, Rosa P (1998). "Borrelia burgdorferi Erp proteins are immunogenic in mammals infected by tick bite, and their synthesis is inducible in cultured bacteria". Infect. Immun. 66 (6): 2648–54. PMC 108251. PMID 9596729.
- Stevenson B, El-Hage N, Hines MA, Miller JC, Babb K (2002). "Differential binding of host complement inhibitor factor H by Borrelia burgdorferi Erp surface proteins: a possible mechanism underlying the expansive host range of Lyme disease spirochetes". Infect. Immun. 70 (2): 491–7. doi:10.1128/IAI.70.2.491-497.2002. PMC 127719. PMID 11796574.
- Woodman ME, Cooley AE, Miller JC, Lazarus JJ, Tucker K, Bykowski T, Botto M, Hellwage J, Wooten RM, Stevenson B (2007). "Borrelia burgdorferi binding of host complement regulator factor H is not required for efficient mammalian infection". Infect. Immun. 75 (6): 3131–9. doi:10.1128/IAI.01923-06. PMC 1932899. PMID 17420242.
- Kullberg, Bart Jan; Vrijmoeth, Hedwig D; van de Schoor, Freek; Hovius, Joppe W (26 May 2020). "Lyme borreliosis: diagnosis and management". BMJ: m1041. doi:10.1136/bmj.m1041.
|access-date=requires|url=(help) - Embers ME, Ramamoorthy R, Philipp MT (2004). "Survival strategies of Borrelia burgdorferi, the etiologic agent of Lyme disease". Microbes Infect. 6 (3): 312–318. doi:10.1016/j.micinf.2003.11.014. PMID 15065567.
- Liang FT, Yan J, Mbow ML, et al. (2004). "Borrelia burgdorferi changes its surface antigenic expression in response to host immune responses". Infect Immun. 72 (10): 5759–5767. doi:10.1128/IAI.72.10.5759-5767.2004. PMC 517580. PMID 15385475.
- Gilmore RD, Howison RR, Schmit VL, et al. (2007). "Temporal expression analysis of the Borrelia burgdorferi paralogous gene family 54 genes BBA64, BBA65, and BBA66 during persistent infection in mice". Infect. Immun. 75 (6): 2753–2764. doi:10.1128/IAI.00037-07. PMC 1932849. PMID 17371862.
External links
- Atlas of Borrelia (images of spirochetal, spheroplast and granular forms)
- NCBI Taxonomy Browser – Borrelia
- Borrelia burgdorferi B31 Genome Page
- Borrelia garinii PBi Genome Page
- Borrelia afzelli PKo Genome Page
- Schwan TG, Piesman J (February 2002). "Vector interactions and molecular adaptations of lyme disease and relapsing fever spirochetes associated with transmission by ticks". Emerging Infect. Dis. 8 (2): 115–21. doi:10.3201/eid0802.010198. PMC 2732444. PMID 11897061.