IRS1
Insulin receptor substrate 1 (IRS-1) is a signaling adapter protein that in humans is encoded by the IRS-1 gene.[5] It is a 131 kDa protein with amino acid sequence of 1242 residues.[6] It contains a single pleckstrin homology (PH) domain at the N-terminus and a PTB domain ca. 40 residues downstream of this, followed by a poorly conserved C-terminus tail.[7] Together with IRS2, IRS3 (pseudogene) and IRS4, it is homologous to the Drosophila protein chico, whose disruption extends the median lifespan of flies up to 48%.[8] Similarly, Irs1 mutant mice experience moderate life extension and delayed age-related pathologies.[9]




Function
Insulin receptor substrate 1 plays a key role in transmitting signals from the insulin and insulin-like growth factor-1 (IGF-1) receptors to intracellular pathways PI3K / Akt and Erk MAP kinase pathways. Tyrosine phosphorylation of IRS-1 by insulin receptor (IR) introduces multiple binding sites for proteins bearing SH2 homology domain, such as PI3K, Grb-2/Sos complex and SHP2. PI3K, involved in interaction with IRS-1, produces PIP3, which, in turn, recruits Akt kinase. Further, Akt kinase is activated via phosphorylation of its T308 residue and analogous sites in PKC by PDK1. This phosphorylation is absent in tissues lacking IRS-1. The cascade is followed by glucose uptake. Formation of the Grb-2/Sos complex, also known as the RAS guanine nucleotide exchange factor complex, results in ERK1/2 activation. IRS-1 signal transduction may be inhibited by SHP2 in some tissues.[7]
Tyrosine phosphorylation of the insulin receptors or IGF-1 receptors, upon extracellular ligand binding, induces the cytoplasmic binding of IRS-1 to these receptors, through its PTB domains. Multiple tyrosine residues of IRS-1 itself are then phosphorylated by these receptors. This enables IRS-1 to activate several signalling pathways, including the PI3K pathway and the MAP kinase pathway.
An alternative multi-site phosphorylation of Serine/Tyrosine in IRS-1 regulates insulin signaling positively and negatively. C-terminal region contains most of the phosphorylation sites of the protein. The C-terminal tail is not structured, therefore the mechanisms of regulation of IRS-1 by phosphorylation still remain unclear. It has been shown that TNFα causes insulin resistance and multi-site S/T phosphorylation, which results in block of interaction between IRS-1 and juxtamembrane domain peptide, thus converting IRS-1 into an inactive state.[7]
IRS-1 plays important biological function for both metabolic and mitogenic (growth promoting) pathways: mice deficient of IRS1 have only a mild diabetic phenotype, but a pronounced growth impairment, i.e., IRS-1 knockout mice only reach 50% of the weight of normal mice.
Regulation
The cellular protein levels of IRS-1 are regulated by the Cullin7 E3 ubiquitin ligase, which targets IRS-1 for ubiquitin mediated degradation by the proteasome.[10] Different Serine phosphorylation of IRS-1, caused by various molecules, such as fatty acids, TNFα and AMPK, has different effects on the protein, but most of these effects include cellular re-localization, conformational and steric changes. These processes lead to decrease in Tyrosine phosphorylation by insulin receptors and diminished PI3K recruitment. Altogether, these mechanisms stimulate IRS-1 degradation and insulin resistance. Other inhibitory pathways include SOCS proteins and O-GlcNAcylation of IRS-1. SOCS proteins act by binding to IR and by interfering with IR phosphorylation of IRS-1, therefore attenuating insulin signaling. They can also bind to JAK, causing a subsequent decrease in IRS-1 tyrosine phosphorylation. During insulin resistance induced by hyperglycemia, glucose accumulates in tissues as its hexosamine metabolite UDP-GlcNAc. This metabolite if present in high amounts leads to O-GlcNAc protein modifications. IRS-1 can undergo this modification, which results in its phosphorylation and functional suppression.[11]
Interactions
IRS1 has been shown to interact with:
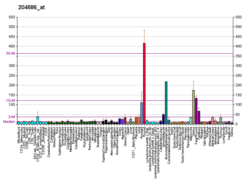
Role in cancer
IRS-1, as a signalling adapter protein, is able to integrate different signalling cascades, which indicates its possible role in cancer progression.[36] IRS-1 protein is known to be involved in various types of cancer, including colorectal,[37] lung,[38] prostate and breast cancer.[39] IRS-1 integrates signalling from insulin receptor (InsR), insulin-like growth factor-1 receptor (IGF1R) and many other cytokine receptors and is elevated in β-catenin induced cells. Some evidence shows that TCF/LEF-β-catenin complexes directly regulate IRS-1. IRS-1 is required for maintenance of neoplasmic phenotype in adenomatous polyposis coli (APC) - mutated cells, it is also needed for transformation in ectopically expressing oncogenic β-catenin cells. IRS-1 dominant-negative mutant functions as tumor suppressor, whereas ectopic IRS-1 stimulates oncogenic transformation. IRS-1 is upregulated in colorectal cancers (CRC) with elevated levels of β-catenin, c-MYC, InsRβ and IGF1R. IRS-1 promotes CRC metastasis to the liver.[37] Decreased apoptosis of crypt stem cells is associated with colon cancer risk. Reduced expression of IRS-1 in Apc (min/+) mutated mice showes increased irradiation-induced apoptosis in crypt. Deficiency in IRS-1 - partial (+/-) or absolute (-/-) - in Apc (min/+) mice demonstrates reduced amount of tumors comparing to IRS-1 (+/+)/ Apc (min/+) mice.[40]
In lung adenocarcinoma cell line A549 overexpression of IRS-1 leads to reduced growth. Tumor infiltrating neutrophils have recently been thought to adjust tumor growth and invasiveness. Neutrophil elastase is shown to degrade IRS-1 by gaining access to endosomal compartment of carcinoma cell. IRS-1 degradation induces cell proliferation in mouse and human adenocarcinomas. Ablation of IRS-1 alters downstream signalling through phosphatidylinositol-3 kinase (PI3K), causing an increased interaction of it with platelet-derived growth factor receptor (PDGFR). Therefore, IRS-1 acts as major regulator of PI3K in lung adenocarcinoma.[38]
Some evidence shows role of IRS-1 in hepatocellular carcinoma (HCC). In rat model, IRS-1 focal overexpression is associated with early events of hepatocarcinogenesis. During progression of preneoplastic foci into hepatocellular carcinomas expression of IRS-1 gradually decreases, which is characterises a metabolic shift heading towards malignant neoplastic phenotype.[41] Transgenic mice, co-expressing IRS-1 and hepatitis Bx (HBx) protein, demonstrate higher rate of hepatocellular displasia that results in HCC development. Expressed alone, IRS-1 and HBx are not sufficient to induce neoplastic alterations in the liver, though their paired expression switches on IN/IRS-1/MAPK and Wnt/β-catenin cascades, causing HCC transformation.[42]
LNCaP prostate cancer cells increase cell adhesion and diminish cell motility via IGF-1 independent mechanism, when IRS-1 is ectopically expressed in the cells. These effects are mediated by PI3K. Uncanonical phosphorylation of Serine 612 by PI3K of IRS-1 protein is due to hyper-activation of Akt/PKB pathway in LNCaP. IRS-1 interacts with integrin α5β1, activating an alternative signalling cascade. This cascade results in decreased cell motility opposing to IGF-1 - dependent mechanism. Loss of IRS-1 expression and PTEN mutations in LNCaP cells could promote metastasis.[43] Ex vivo studies of IRS-1 involvement in prostate cancer show ambiguous results. Down-regulation of IGF1R in bone marrow biopsies of metastatic prostate cancer goes along with down-regulation of IRS-1 and significant reduction of PTEN in 3 out of 12 cases. Most of the tumors still express IRS-1 and IGF1R during progression of the metastatic disease.[44]
IRS-1 has a functional role in breast cancer progression and metastasis. Overexpression of PTEN in MCF-7 epithelial breast cancer cells inhibits cell growth by inhibiting MAPK pathway. ERK phosphorylation through IRS-1/Grb-2/Sos pathway is inhibited by phosphatase activity of PTEN. PTEN does not have effect on IRS-1 independent MAPK activation. When treated with insulin, ectopic expression of PTEN in MCF-7 suppresses IRS-1/Grb-2/Sos complex formation due to differential phosphorylation of IRS-1.[45] Overexpression of IRS-1 has been linked to antiestrogen resistance and hormone independence in breast cancer. Tamoxifen (TAM) inhibits IRS-1 function, therefore suppressing IRS-1/PI3K signalling cascade in estrogen receptor positive (ER+) MCF-7 cell line. IRS-1 siRNA is able to reduce IRS-1 transcript level, thereby reducing protein expression in MCF-7 ER+ cells. Reduction of IRS-1 leads to decreased survival of these cells. siRNA treatment effects are additive to effects of TAM treatment.[46] IGFRs and estrogen coaction facilitates growth in different breast cancer cell lines, however amplification of IGF1R signalling can abrogate need of estrogen for transformation and growth of MCF-7 cells. IRS-1 overexpression in breast cancer cells decreased estrogen requirements. This decrease is dependent on IRS-1 levels in the cells.[47] Estradiol enhances expression of IRS-1 and activity of ERK1/2 and PI3K/Akt pathways in MCF-7 and CHO cells transfected with mouse IRS-1 promoter. Estradiol acts directly on IRS-1 regulatory sequences and positively regulates IRS-1 mRNA production.[48] Decreased anchorage- dependent/independent cell growth and initiation of cell death under low growth factor and estrogen conditions are observed in MCF-7 cells with down-regulated IRS-1.[49] mir126 is underexpressed in breast cancer cells. mir126 targets IRS-1 at transcriptional level and inhibits transition from G1/G0 phase to S phase during cell cycle in HEK293 and MCF-7 cells.[50] Transgenic mice overexpressing IRS-1 develop metastatic breast cancer.The tumors demonstrate squamous differentiation which is associated with β-catenin pathway. IRS-1 interacts with β-catenin both in vitro and in vivo.[51] IRS-1 and its homologue IRS-2 play distinct roles in breast cancer progression and metastasis. Overexpression of either one is sufficient to cause tumorogenesis in vivo. Frequency of lung metastasis in IRS-1 deficient tumor is elevated opposing to IRS-2 deficient tumor, where it is decreased. Basically, IRS-2 has a positive impact on metastasis of breast cancer whereas a stronger metastatic potential is observed when IRS-1 is down-regulated. IRS-1 is strongly expressed in ductal carcinoma in situ, when IRS-2 is elevated in invasive tumors. Increased IRS-1 makes MCF-7 cells susceptible to specific chemotherapeutic agents, such as taxol, etoposide, and vincristine.Therefore, IRS-1 can be a good pointer of specific drug therapies effectiveness for breast cancer treatment.[52]
References
- GRCh38: Ensembl release 89: ENSG00000169047 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000055980 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Sun XJ, Rothenberg P, Kahn CR, Backer JM, Araki E, Wilden PA, Cahill DA, Goldstein BJ, White MF (July 1991). "Structure of the insulin receptor substrate IRS-1 defines a unique signal transduction protein". Nature. 352 (6330): 73–7. doi:10.1038/352073a0. PMID 1648180.
- "IRS1 - Insulin receptor substrate 1 - Homo sapiens (Human) - IRS1 gene & protein". www.uniprot.org. Retrieved 2016-04-21.
- Copps KD, White MF (October 2012). "Regulation of insulin sensitivity by serine/threonine phosphorylation of insulin receptor substrate proteins IRS1 and IRS2". Diabetologia. 55 (10): 2565–82. doi:10.1007/s00125-012-2644-8. PMC 4011499. PMID 22869320.
- Clancy DJ, Gems D, Harshman LG, Oldham S, Stocker H, Hafen E, Leevers SJ, Partridge L (April 2001). "Extension of life-span by loss of CHICO, a Drosophila insulin receptor substrate protein". Science. 292 (5514): 104–6. doi:10.1126/science.1057991. PMID 11292874.
- Selman C, Lingard S, Choudhury AI, Batterham RL, Claret M, Clements M, Ramadani F, Okkenhaug K, Schuster E, Blanc E, Piper MD, Al-Qassab H, Speakman JR, Carmignac D, Robinson IC, Thornton JM, Gems D, Partridge L, Withers DJ (March 2008). "Evidence for lifespan extension and delayed age-related biomarkers in insulin receptor substrate 1 null mice". FASEB Journal. 22 (3): 807–18. doi:10.1096/fj.07-9261com. PMID 17928362.
- Xu X, Sarikas A, Dias-Santagata DC, Dolios G, Lafontant PJ, Tsai SC, Zhu W, Nakajima H, Nakajima HO, Field LJ, Wang R, Pan ZQ (May 2008). "The CUL7 E3 ubiquitin ligase targets insulin receptor substrate 1 for ubiquitin-dependent degradation". Molecular Cell. 30 (4): 403–14. doi:10.1016/j.molcel.2008.03.009. PMC 2633441. PMID 18498745.
- Gual P, Le Marchand-Brustel Y, Tanti JF (January 2005). "Positive and negative regulation of insulin signaling through IRS-1 phosphorylation". Biochimie. 87 (1): 99–109. doi:10.1016/j.biochi.2004.10.019. PMID 15733744.
- Ueno H, Kondo E, Yamamoto-Honda R, Tobe K, Nakamoto T, Sasaki K, Mitani K, Furusaka A, Tanaka T, Tsujimoto Y, Kadowaki T, Hirai H (February 2000). "Association of insulin receptor substrate proteins with Bcl-2 and their effects on its phosphorylation and antiapoptotic function". Molecular Biology of the Cell. 11 (2): 735–46. doi:10.1091/mbc.11.2.735. PMC 14806. PMID 10679027.
- Skolnik EY, Lee CH, Batzer A, Vicentini LM, Zhou M, Daly R, Myers MJ, Backer JM, Ullrich A, White MF (May 1993). "The SH2/SH3 domain-containing protein GRB2 interacts with tyrosine-phosphorylated IRS1 and Shc: implications for insulin control of ras signalling". The EMBO Journal. 12 (5): 1929–36. doi:10.1002/j.1460-2075.1993.tb05842.x. PMC 413414. PMID 8491186.
- Morrison KB, Tognon CE, Garnett MJ, Deal C, Sorensen PH (August 2002). "ETV6-NTRK3 transformation requires insulin-like growth factor 1 receptor signaling and is associated with constitutive IRS-1 tyrosine phosphorylation". Oncogene. 21 (37): 5684–95. doi:10.1038/sj.onc.1205669. PMID 12173038.
- Giorgetti-Peraldi S, Peyrade F, Baron V, Van Obberghen E (December 1995). "Involvement of Janus kinases in the insulin signaling pathway". European Journal of Biochemistry / FEBS. 234 (2): 656–60. doi:10.1111/j.1432-1033.1995.656_b.x. PMID 8536716.
- Aguirre V, Werner ED, Giraud J, Lee YH, Shoelson SE, White MF (January 2002). "Phosphorylation of Ser307 in insulin receptor substrate-1 blocks interactions with the insulin receptor and inhibits insulin action". The Journal of Biological Chemistry. 277 (2): 1531–7. doi:10.1074/jbc.M101521200. PMID 11606564.
- Sawka-Verhelle D, Tartare-Deckert S, White MF, Van Obberghen E (March 1996). "Insulin receptor substrate-2 binds to the insulin receptor through its phosphotyrosine-binding domain and through a newly identified domain comprising amino acids 591-786". The Journal of Biological Chemistry. 271 (11): 5980–3. doi:10.1074/jbc.271.11.5980. PMID 8626379.
- Tartare-Deckert S, Sawka-Verhelle D, Murdaca J, Van Obberghen E (October 1995). "Evidence for a differential interaction of SHC and the insulin receptor substrate-1 (IRS-1) with the insulin-like growth factor-I (IGF-I) receptor in the yeast two-hybrid system". The Journal of Biological Chemistry. 270 (40): 23456–60. doi:10.1074/jbc.270.40.23456. PMID 7559507.
- Dey BR, Frick K, Lopaczynski W, Nissley SP, Furlanetto RW (June 1996). "Evidence for the direct interaction of the insulin-like growth factor I receptor with IRS-1, Shc, and Grb10". Molecular Endocrinology. 10 (6): 631–41. doi:10.1210/mend.10.6.8776723. PMID 8776723.
- Mañes S, Mira E, Gómez-Mouton C, Zhao ZJ, Lacalle RA, Martínez-A C (April 1999). "Concerted activity of tyrosine phosphatase SHP-2 and focal adhesion kinase in regulation of cell motility". Molecular and Cellular Biology. 19 (4): 3125–35. doi:10.1128/mcb.19.4.3125. PMC 84106. PMID 10082579.
- Gual P, Baron V, Lequoy V, Van Obberghen E (March 1998). "Interaction of Janus kinases JAK-1 and JAK-2 with the insulin receptor and the insulin-like growth factor-1 receptor". Endocrinology. 139 (3): 884–93. doi:10.1210/endo.139.3.5829. PMID 9492017.
- Johnston JA, Wang LM, Hanson EP, Sun XJ, White MF, Oakes SA, Pierce JH, O'Shea JJ (December 1995). "Interleukins 2, 4, 7, and 15 stimulate tyrosine phosphorylation of insulin receptor substrates 1 and 2 in T cells. Potential role of JAK kinases". The Journal of Biological Chemistry. 270 (48): 28527–30. doi:10.1074/jbc.270.48.28527. PMID 7499365.
- Kawazoe Y, Naka T, Fujimoto M, Kohzaki H, Morita Y, Narazaki M, Okumura K, Saitoh H, Nakagawa R, Uchiyama Y, Akira S, Kishimoto T (January 2001). "Signal transducer and activator of transcription (STAT)-induced STAT inhibitor 1 (SSI-1)/suppressor of cytokine signaling 1 (SOCS1) inhibits insulin signal transduction pathway through modulating insulin receptor substrate 1 (IRS-1) phosphorylation". The Journal of Experimental Medicine. 193 (2): 263–9. doi:10.1084/jem.193.2.263. PMC 2193341. PMID 11208867.
- Aguirre V, Uchida T, Yenush L, Davis R, White MF (March 2000). "The c-Jun NH(2)-terminal kinase promotes insulin resistance during association with insulin receptor substrate-1 and phosphorylation of Ser(307)". The Journal of Biological Chemistry. 275 (12): 9047–54. doi:10.1074/jbc.275.12.9047. PMID 10722755.
- Hadari YR, Tzahar E, Nadiv O, Rothenberg P, Roberts CT, LeRoith D, Yarden Y, Zick Y (September 1992). "Insulin and insulinomimetic agents induce activation of phosphatidylinositol 3'-kinase upon its association with pp185 (IRS-1) in intact rat livers". The Journal of Biological Chemistry. 267 (25): 17483–6. PMID 1381348.
- Gual P, Gonzalez T, Grémeaux T, Barres R, Le Marchand-Brustel Y, Tanti JF (July 2003). "Hyperosmotic stress inhibits insulin receptor substrate-1 function by distinct mechanisms in 3T3-L1 adipocytes". The Journal of Biological Chemistry. 278 (29): 26550–7. doi:10.1074/jbc.M212273200. PMID 12730242.
- Hamer I, Foti M, Emkey R, Cordier-Bussat M, Philippe J, De Meyts P, Maeder C, Kahn CR, Carpentier JL (May 2002). "An arginine to cysteine(252) mutation in insulin receptors from a patient with severe insulin resistance inhibits receptor internalisation but preserves signalling events". Diabetologia. 45 (5): 657–67. doi:10.1007/s00125-002-0798-5. PMID 12107746.
- Xia X, Serrero G (August 1999). "Multiple forms of p55PIK, a regulatory subunit of phosphoinositide 3-kinase, are generated by alternative initiation of translation". The Biochemical Journal. 341 (3): 831–7. doi:10.1042/0264-6021:3410831. PMC 1220424. PMID 10417350.
- Mothe I, Delahaye L, Filloux C, Pons S, White MF, Van Obberghen E (December 1997). "Interaction of wild type and dominant-negative p55PIK regulatory subunit of phosphatidylinositol 3-kinase with insulin-like growth factor-1 signaling proteins". Molecular Endocrinology. 11 (13): 1911–23. doi:10.1210/mend.11.13.0029. PMID 9415396.
- Lebrun P, Mothe-Satney I, Delahaye L, Van Obberghen E, Baron V (November 1998). "Insulin receptor substrate-1 as a signaling molecule for focal adhesion kinase pp125(FAK) and pp60(src)". The Journal of Biological Chemistry. 273 (48): 32244–53. doi:10.1074/jbc.273.48.32244. PMID 9822703.
- Kuhné MR, Pawson T, Lienhard GE, Feng GS (June 1993). "The insulin receptor substrate 1 associates with the SH2-containing phosphotyrosine phosphatase Syp". The Journal of Biological Chemistry. 268 (16): 11479–81. PMID 8505282.
- Myers MG, Mendez R, Shi P, Pierce JH, Rhoads R, White MF (October 1998). "The COOH-terminal tyrosine phosphorylation sites on IRS-1 bind SHP-2 and negatively regulate insulin signaling". The Journal of Biological Chemistry. 273 (41): 26908–14. doi:10.1074/jbc.273.41.26908. PMID 9756938.
- Goldstein BJ, Bittner-Kowalczyk A, White MF, Harbeck M (February 2000). "Tyrosine dephosphorylation and deactivation of insulin receptor substrate-1 by protein-tyrosine phosphatase 1B. Possible facilitation by the formation of a ternary complex with the Grb2 adaptor protein". The Journal of Biological Chemistry. 275 (6): 4283–9. doi:10.1074/jbc.275.6.4283. PMID 10660596.
- Ravichandran LV, Chen H, Li Y, Quon MJ (October 2001). "Phosphorylation of PTP1B at Ser(50) by Akt impairs its ability to dephosphorylate the insulin receptor". Molecular Endocrinology. 15 (10): 1768–80. doi:10.1210/mend.15.10.0711. PMID 11579209.
- Craparo A, Freund R, Gustafson TA (April 1997). "14-3-3 (epsilon) interacts with the insulin-like growth factor I receptor and insulin receptor substrate I in a phosphoserine-dependent manner". The Journal of Biological Chemistry. 272 (17): 11663–9. doi:10.1074/jbc.272.17.11663. PMID 9111084.
- Dearth RK, Cui X, Kim HJ, Hadsell DL, Lee AV (March 2007). "Oncogenic transformation by the signaling adaptor proteins insulin receptor substrate (IRS)-1 and IRS-2". Cell Cycle. 6 (6): 705–13. doi:10.4161/cc.6.6.4035. PMID 17374994.
- Esposito DL, Aru F, Lattanzio R, Morgano A, Abbondanza M, Malekzadeh R, Bishehsari F, Valanzano R, Russo A, Piantelli M, Moschetta A, Lotti LV, Mariani-Costantini R (2012-04-27). "The insulin receptor substrate 1 (IRS1) in intestinal epithelial differentiation and in colorectal cancer". PLOS ONE. 7 (4): e36190. doi:10.1371/journal.pone.0036190. PMC 3338610. PMID 22558377.
- Houghton AM, Rzymkiewicz DM, Ji H, Gregory AD, Egea EE, Metz HE, Stolz DB, Land SR, Marconcini LA, Kliment CR, Jenkins KM, Beaulieu KA, Mouded M, Frank SJ, Wong KK, Shapiro SD (February 2010). "Neutrophil elastase-mediated degradation of IRS-1 accelerates lung tumor growth". Nature Medicine. 16 (2): 219–23. doi:10.1038/nm.2084. PMC 2821801. PMID 20081861.
- Gibson SL, Ma Z, Shaw LM (March 2007). "Divergent roles for IRS-1 and IRS-2 in breast cancer metastasis". Cell Cycle. 6 (6): 631–7. doi:10.4161/cc.6.6.3987. PMID 17361103.
- Ramocki NM, Wilkins HR, Magness ST, Simmons JG, Scull BP, Lee GH, McNaughton KK, Lund PK (January 2008). "Insulin receptor substrate-1 deficiency promotes apoptosis in the putative intestinal crypt stem cell region, limits Apcmin/+ tumors, and regulates Sox9". Endocrinology. 149 (1): 261–7. doi:10.1210/en.2007-0869. PMC 2194604. PMID 17916629.
- Nehrbass D, Klimek F, Bannasch P (February 1998). "Overexpression of insulin receptor substrate-1 emerges early in hepatocarcinogenesis and elicits preneoplastic hepatic glycogenosis". The American Journal of Pathology. 152 (2): 341–5. PMC 1857952. PMID 9466558.
- Longato L, de la Monte S, Kuzushita N, Horimoto M, Rogers AB, Slagle BL, Wands JR (June 2009). "Overexpression of insulin receptor substrate-1 and hepatitis Bx genes causes premalignant alterations in the liver". Hepatology. 49 (6): 1935–43. doi:10.1002/hep.22856. PMC 2754284. PMID 19475691.
- Reiss K, Wang JY, Romano G, Tu X, Peruzzi F, Baserga R (January 2001). "Mechanisms of regulation of cell adhesion and motility by insulin receptor substrate-1 in prostate cancer cells". Oncogene. 20 (4): 490–500. doi:10.1038/sj.onc.1204112. PMID 11313980.
- Hellawell GO, Turner GD, Davies DR, Poulsom R, Brewster SF, Macaulay VM (May 2002). "Expression of the type 1 insulin-like growth factor receptor is up-regulated in primary prostate cancer and commonly persists in metastatic disease". Cancer Research. 62 (10): 2942–50. PMID 12019176.
- Weng LP, Smith WM, Brown JL, Eng C (March 2001). "PTEN inhibits insulin-stimulated MEK/MAPK activation and cell growth by blocking IRS-1 phosphorylation and IRS-1/Grb-2/Sos complex formation in a breast cancer model". Human Molecular Genetics. 10 (6): 605–16. doi:10.1093/hmg/10.6.605. PMID 11230180.
- Cesarone G, Edupuganti OP, Chen CP, Wickstrom E (2007-12-01). "Insulin receptor substrate 1 knockdown in human MCF7 ER+ breast cancer cells by nuclease-resistant IRS1 siRNA conjugated to a disulfide-bridged D-peptide analogue of insulin-like growth factor 1". Bioconjugate Chemistry. 18 (6): 1831–40. doi:10.1021/bc070135v. PMID 17922544.
- Surmacz E, Burgaud JL (November 1995). "Overexpression of insulin receptor substrate 1 (IRS-1) in the human breast cancer cell line MCF-7 induces loss of estrogen requirements for growth and transformation". Clinical Cancer Research. 1 (11): 1429–36. PMID 9815941.
- Mauro L, Salerno M, Panno ML, Bellizzi D, Sisci D, Miglietta A, Surmacz E, Andò S (November 2001). "Estradiol increases IRS-1 gene expression and insulin signaling in breast cancer cells". Biochemical and Biophysical Research Communications. 288 (3): 685–9. doi:10.1006/bbrc.2001.5815. PMID 11676497.
- Nolan MK, Jankowska L, Prisco M, Xu S, Guvakova MA, Surmacz E (September 1997). "Differential roles of IRS-1 and SHC signaling pathways in breast cancer cells". International Journal of Cancer. 72 (5): 828–34. doi:10.1002/(sici)1097-0215(19970904)72:5<828::aid-ijc20>3.0.co;2-3. PMID 9311601.
- Zhang J, Du YY, Lin YF, Chen YT, Yang L, Wang HJ, Ma D (December 2008). "The cell growth suppressor, mir-126, targets IRS-1". Biochemical and Biophysical Research Communications. 377 (1): 136–40. doi:10.1016/j.bbrc.2008.09.089. PMID 18834857.
- Dearth RK, Cui X, Kim HJ, Kuiatse I, Lawrence NA, Zhang X, Divisova J, Britton OL, Mohsin S, Allred DC, Hadsell DL, Lee AV (December 2006). "Mammary tumorigenesis and metastasis caused by overexpression of insulin receptor substrate 1 (IRS-1) or IRS-2". Molecular and Cellular Biology. 26 (24): 9302–14. doi:10.1128/MCB.00260-06. PMC 1698542. PMID 17030631.
- Porter HA, Perry A, Kingsley C, Tran NL, Keegan AD (September 2013). "IRS1 is highly expressed in localized breast tumors and regulates the sensitivity of breast cancer cells to chemotherapy, while IRS2 is highly expressed in invasive breast tumors". Cancer Letters. 338 (2): 239–48. doi:10.1016/j.canlet.2013.03.030. PMC 3761875. PMID 23562473.
Further reading
- Jiang H, Harris MB, Rothman P (June 2000). "IL-4/IL-13 signaling beyond JAK/STAT". The Journal of Allergy and Clinical Immunology. 105 (6 Pt 1): 1063–70. doi:10.1067/mai.2000.107604. PMID 10856136.
- Bezerra RM, Chadid TT, Altemani CM, Sales TS, Menezes R, Soares MC, Saad ST, Saad MJ (February 2004). "Lack of Arg972 polymorphism in the IRS1 gene in Parakanã Brazilian Indians". Human Biology. 76 (1): 147–51. doi:10.1353/hub.2004.0015. PMID 15222685.
- Gibson SL, Ma Z, Shaw LM (March 2007). "Divergent roles for IRS-1 and IRS-2 in breast cancer metastasis". Cell Cycle. 6 (6): 631–7. doi:10.4161/cc.6.6.3987. PMID 17361103.
- Dearth RK, Cui X, Kim HJ, Hadsell DL, Lee AV (March 2007). "Oncogenic transformation by the signaling adaptor proteins insulin receptor substrate (IRS)-1 and IRS-2". Cell Cycle. 6 (6): 705–13. doi:10.4161/cc.6.6.4035. PMID 17374994.
PDB gallery | |
|---|---|
|