Gonadotropin-releasing hormone
Gonadotropin-releasing hormone (GnRH) is a releasing hormone responsible for the release of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) from the anterior pituitary. GnRH is a tropic peptide hormone synthesized and released from GnRH neurons within the hypothalamus. The peptide belongs to gonadotropin-releasing hormone family. It constitutes the initial step in the hypothalamic–pituitary–gonadal axis.
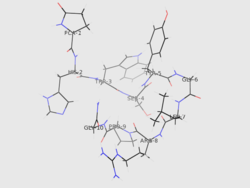


Structure
The identity[5] of GnRH was clarified by the 1977 Nobel Laureates Roger Guillemin and Andrew V. Schally:[6]
As is standard for peptide representation, the sequence is given from amino terminus to carboxyl terminus; also standard is omission of the designation of chirality, with assumption that all amino acids are in their L- form. The abbreviations are the standard abbreviations for the corresponding proteinogenic amino acids, except for pyroGlu, which refers to pyroglutamic acid, a derivative of glutamic acid. The NH2 at the carboxyl terminus indicates that rather than terminating as a free carboxylate, it terminates as a carboxamide.
Synthesis
The gene, GNRH1, for the GnRH precursor is located on chromosome 8. In mammals, the linear decapeptide end-product is synthesized from an 89-amino acid preprohormone in the preoptic anterior hypothalamus. It is the target of various regulatory mechanisms of the hypothalamic–pituitary–gonadal axis, such as being inhibited by increased estrogen levels in the body.
Function
GnRH is secreted in the hypophysial portal bloodstream at the median eminence.[7] The portal blood carries the GnRH to the pituitary gland, which contains the gonadotrope cells, where GnRH activates its own receptor, gonadotropin-releasing hormone receptor (GnRHR), a seven-transmembrane G-protein-coupled receptor that stimulates the beta isoform of Phosphoinositide phospholipase C, which goes on to mobilize calcium and protein kinase C. This results in the activation of proteins involved in the synthesis and secretion of the gonadotropins LH and FSH. GnRH is degraded by proteolysis within a few minutes.
GnRH activity is very low during childhood, and is activated at puberty or adolescence. During the reproductive years, pulse activity is critical for successful reproductive function as controlled by feedback loops. However, once a pregnancy is established, GnRH activity is not required. Pulsatile activity can be disrupted by hypothalamic-pituitary disease, either dysfunction (i.e., hypothalamic suppression) or organic lesions (trauma, tumor). Elevated prolactin levels decrease GnRH activity. In contrast, hyperinsulinemia increases pulse activity leading to disorderly LH and FSH activity, as seen in polycystic ovary syndrome (PCOS). GnRH formation is congenitally absent in Kallmann syndrome.
Control of FSH and LH
At the pituitary, GnRH stimulates the synthesis and secretion of follicle-stimulating hormone (FSH) and luteinizing hormone (LH).[8] These processes are controlled by the size and frequency of GnRH pulses, as well as by feedback from androgens and estrogens. Low-frequency GnRH pulses are required for FSH release, whereas high-frequency GnRH pulses stimulate LH pulses in a one-to-one manner.[9]
There are differences in GnRH secretion between females and males. In males, GnRH is secreted in pulses at a constant frequency; however, in females, the frequency of the pulses varies during the menstrual cycle, and there is a large surge of GnRH just before ovulation.[10]
GnRH secretion is pulsatile in all vertebrates,[11] and is necessary for correct reproductive function. Thus, a single hormone, GnRH1, controls a complex process of follicular growth, ovulation, and corpus luteum maintenance in the female, and spermatogenesis in the male.
Neurohormone
GnRH is considered a neurohormone, a hormone produced in a specific neural cell and released at its neural terminal. A key area for production of GnRH is the preoptic area of the hypothalamus, which contains most of the GnRH-secreting neurons. GnRH neurons originate in the nose and migrate into the brain, where they are scattered throughout the medial septum and hypothalamus and connected by very long >1-millimeter-long dendrites. These bundle together so they receive shared synaptic input, a process that allows them to synchronize their GnRH release.[7]
The GnRH neurons are regulated by many different afferent neurons, using several different transmitters (including norepinephrine, GABA, glutamate). For instance, dopamine appears to stimulate LH release (through GnRH) in estrogen-progesterone-primed females; dopamine may inhibit LH release in ovariectomized females.[8] Kisspeptin appears to be an important regulator of GnRH release.[12] GnRH release can also be regulated by estrogen. It has been reported that there are kisspeptin-producing neurons that also express estrogen receptor alpha.[13]
Other organs
GnRH is found in organs outside of the hypothalamus and pituitary, and its role in other life processes is poorly understood. For instance, there is likely to be a role for GnRH1 in the placenta and in the gonads. GnRH and GnRH receptors are also found in cancers of the breast, ovary, prostate, and endometrium.[14]
Effects of behavior
GnRH production/release is one of the few confirmed examples of behavior influencing hormones, rather than the other way around. Cichlid fish that become socially dominant in turn experience an upregulation of GnRH secretion whereas cichlid fish that are socially subordinate have a down regulation of GnRH secretion.[15] Besides secretion, the social environment as well as their behavior affects the size of GnRH neurons. Specifically, males that are more territorial have larger GnRH neurons than males that are less territorial. Differences are also seen in females, with brooding females having smaller GnRH neurons than either spawning or control females.[16] These examples suggest that GnRH is a socially regulated hormone.
Medical uses
Natural GnRH was previously prescribed as gonadorelin hydrochloride (Factrel)[17] and gonadorelin diacetate tetrahydrate (Cystorelin)[18] for use in treating human diseases. Modifications of the decapeptide structure of GnRH to increase half life have led to GnRH1 analog medications that either stimulate (GnRH1 agonists) or suppress (GnRH antagonists) the gonadotropins. These synthetic analogs have replaced the natural hormone in clinical use.
Its analogue leuprorelin is used for continuous infusion, to treat breast cancer, endometriosis, prostate cancer, and following research in the 1980s by researchers, including Dr. Florence Comite of Yale University, it was used to treat precocious puberty.[19][20]
A Cochrane Review is available which investigates whether GnRH analogues, given before or alongside chemotherapy, could prevent damage to women's ovaries caused by chemotherapy.[21] GnRH agonists appear to be effective in protecting the ovaries during chemotherapy, in terms of menstruation recovery or maintenance, premature ovarian failure and ovulation.
Animal sexual behavior
GnRH activity influences a variety of sexual behaviors. Increased levels of GnRH facilitate sexual displays and behavior in females. GnRH injections enhance copulation solicitation (a type of courtship display) in white-crowned sparrows.[22] In mammals, GnRH injections facilitate sexual behavior of female display behaviors as shown with the musk shrew’s (Suncus murinus) reduced latency in displaying rump presents and tail wagging towards males.[23]
An elevation of GnRH raises males’ testosterone capacity beyond a male's natural testosterone level. Injections of GnRH in male birds immediately after an aggressive territorial encounter results in higher testosterone levels than what is observed naturally during an aggressive territorial encounter.[24]
A compromised GnRH system has aversive effects on reproductive physiology and maternal behavior. In comparison to female mice with a normal GnRH system, female mice with a 30% decrease in GnRH neurons are poor caregivers to their offspring. These mice are more likely to leave their pups scattered rather than grouped together, and will take significantly longer to retrieve their pups.[25]
Veterinary use
The natural hormone is also used in veterinary medicine as a treatment for cattle with cystic ovarian disease. The synthetic analogue deslorelin is used in veterinary reproductive control through a sustained-release implant.
Other names
As with many hormones, GnRH has been called by various names in the medical literature over the decades since its existence was first inferred. They are as follows:
- Gonadotropin-releasing factor (GnRF, GRF); Gonadotropin-releasing hormone (GnRH, GRH)
- Follicle-stimulating hormone-releasing factor (FRF, FSH-RF); Follicle-stimulating hormone-releasing hormone (FRH, FSH-RH)
- Luteinizing hormone-releasing factor (LRF, LHRF); Luteinizing hormone-releasing hormone (LRH, LHRH)
- Follicle-stimulating hormone and luteinizing hormone–releasing factor (FSH/LH-RF); Follicle-stimulating hormone and luteinizing hormone-releasing hormone (FSH/LH-RH)
- Luteinizing hormone and follicle-stimulating hormone–releasing factor (LH/FSH-RF); Luteinizing hormone and follicle-stimulating hormone-releasing hormone (LH/FSH-RH)
- Gonadorelin (INN for pharmaceutical form)
- Gonadoliberin
See also
References
- GRCh38: Ensembl release 89: ENSG00000147437 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000015812 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Kochman K (2012). "Evolution of gonadotropin-releasing hormone (GnRH) structure and its receptor". Journal of Animal and Feed Sciences. 21 (1): 6.
- "The Nobel Prize in Physiology or Medicine 1977". www.nobelprize.org. Nobel Media AB 2014. Retrieved 24 June 2016.
- Campbell RE, Gaidamaka G, Han SK, Herbison AE (June 2009). "Dendro-dendritic bundling and shared synapses between gonadotropin-releasing hormone neurons". Proceedings of the National Academy of Sciences of the United States of America. 106 (26): 10835–40. doi:10.1073/pnas.0903463106. PMC 2705602. PMID 19541658.
- Brown RM (1994). An introduction to Neuroendocrinology. Cambridge, UK: Cambridge University Press. ISBN 978-0-521-42665-7.
- Jayes FC, Britt JH, Esbenshade KL (April 1997). "Role of gonadotropin-releasing hormone pulse frequency in differential regulation of gonadotropins in the gilt" (PDF). Biology of Reproduction. 56 (4): 1012–9. doi:10.1095/biolreprod56.4.1012. PMID 9096885. Archived from the original (PDF) on 2015-09-23.
- Ehlers K, Halvorson L (2013). "Gonadotropin-releasing Hormone (GnRH) and the GnRH Receptor (GnRHR)". The Global Library of Women's Medicine. doi:10.3843/GLOWM.10285. Retrieved 5 November 2014.
- Tsutsumi R, Webster NJ (17 July 2009). "GnRH pulsatility, the pituitary response and reproductive dysfunction". Endocrine Journal. 56 (6): 729–37. doi:10.1507/endocrj.K09E-185. PMC 4307809. PMID 19609045.
- Dungan HM, Clifton DK, Steiner RA (March 2006). "Minireview: kisspeptin neurons as central processors in the regulation of gonadotropin-releasing hormone secretion". Endocrinology. 147 (3): 1154–8. doi:10.1210/en.2005-1282. PMID 16373418.
- Franceschini I, Lomet D, Cateau M, Delsol G, Tillet Y, Caraty A (July 2006). "Kisspeptin immunoreactive cells of the ovine preoptic area and arcuate nucleus co-express estrogen receptor alpha". Neuroscience Letters. 401 (3): 225–30. doi:10.1016/j.neulet.2006.03.039. PMID 16621281.
- Schally AV (1999). "Luteinizing hormone-releasing hormone analogs: their impact on the control of tumorigenesis". Peptides. 20 (10): 1247–62. doi:10.1016/S0196-9781(99)00130-8. PMID 10573298.
- Chee SS, Espinoza WA, Iwaniuk AN, Pakan JM, Gutiérrez-Ibáñez C, Wylie DR, Hurd PL (January 2013). "Social status, breeding state, and GnRH soma size in convict cichlids (Cryptoheros nigrofasciatus)". Behavioural Brain Research. 237: 318–24. doi:10.1016/j.bbr.2012.09.023. PMID 23000535.
- White SA, Nguyen T, Fernald RD (September 2002). "Social regulation of gonadotropin-releasing hormone" (PDF). The Journal of Experimental Biology. 205 (Pt 17): 2567–81. PMID 12151363.
- Drugs.com Factrel: Consumer Drug Information
- Drugs.com Cystorelin: FDA Professional Drug Information
- Comite F, Cutler GB, Rivier J, Vale WW, Loriaux DL, Crowley WF (December 1981). "Short-term treatment of idiopathic precocious puberty with a long-acting analogue of luteinizing hormone-releasing hormone. A preliminary report". The New England Journal of Medicine. 305 (26): 1546–50. doi:10.1056/NEJM198112243052602. PMID 6458765.
- Sonis WA, Comite F, Pescovitz OH, Hench K, Rahn CW, Cutler GB, Loriaux DL, Klein RP (September 1986). "Biobehavioral aspects of precocious puberty". Journal of the American Academy of Child Psychiatry. 25 (5): 674–9. doi:10.1016/S0002-7138(09)60293-4. PMID 3760417.
- Chen H, Xiao L, Li J, Cui L, Huang W (March 2019). "Adjuvant gonadotropin-releasing hormone analogues for the prevention of chemotherapy-induced premature ovarian failure in premenopausal women". The Cochrane Database of Systematic Reviews. 3: CD008018. doi:10.1002/14651858.cd008018.pub3. PMC 6397718. PMID 30827035.
- Maney DL, Richardson RD, Wingfield JC (August 1997). "Central administration of chicken gonadotropin-releasing hormone-II enhances courtship behavior in a female sparrow". Hormones and Behavior. 32 (1): 11–8. doi:10.1006/hbeh.1997.1399. PMID 9344687.
- Schiml PA, Rissman EF (May 2000). "Effects of gonadotropin-releasing hormones, corticotropin-releasing hormone, and vasopressin on female sexual behavior". Hormones and Behavior. 37 (3): 212–20. doi:10.1006/hbeh.2000.1575. PMID 10868484.
- DeVries MS, Winters CP, Jawor JM (June 2012). "Testosterone elevation and response to gonadotropin-releasing hormone challenge by male northern cardinals (Cardinalis cardinalis) following aggressive behavior". Hormones and Behavior. 62 (1): 99–105. doi:10.1016/j.yhbeh.2012.05.008. PMID 22613708.
- Brooks LR, Le CD, Chung WC, Tsai PS (2012). "Maternal behavior in transgenic mice with reduced fibroblast growth factor receptor function in gonadotropin-releasing hormone neurons". Behavioral and Brain Functions. 8: 47. doi:10.1186/1744-9081-8-47. PMC 3503805. PMID 22950531.
Further reading
- Flanagan CA, Millar RP, Illing N (May 1997). "Advances in understanding gonadotrophin-releasing hormone receptor structure and ligand interactions". Reviews of Reproduction. 2 (2): 113–20. doi:10.1530/ror.0.0020113. PMID 9414473.
- Leung PC, Cheng CK, Zhu XM (April 2003). "Multi-factorial role of GnRH-I and GnRH-II in the human ovary". Molecular and Cellular Endocrinology. 202 (1–2): 145–53. doi:10.1016/S0303-7207(03)00076-5. PMID 12770744.
- Gründker C, Emons G (October 2003). "Role of gonadotropin-releasing hormone (GnRH) in ovarian cancer". Reproductive Biology and Endocrinology. 1: 65. doi:10.1186/1477-7827-1-65. PMC 239893. PMID 14594454.
- Limonta P, Moretti RM, Montagnani Marelli M, Motta M (December 2003). "The biology of gonadotropin hormone-releasing hormone: role in the control of tumor growth and progression in humans". Frontiers in Neuroendocrinology. 24 (4): 279–95. doi:10.1016/j.yfrne.2003.10.003. PMID 14726258.
- Janáky T, Juhász A, Bajusz S, Csernus V, Srkalovic G, Bokser L, Milovanovic S, Redding TW, Rékási Z, Nagy A (February 1992). "Analogues of luteinizing hormone-releasing hormone containing cytotoxic groups". Proceedings of the National Academy of Sciences of the United States of America. 89 (3): 972–6. doi:10.1073/pnas.89.3.972. PMC 48367. PMID 1310542.
- Healey SC, Martin NG, Chenevix-Trench G (November 1991). "NcoI RFLP of the human LHRH gene on chromosome 8p". Nucleic Acids Research. 19 (21): 6059. doi:10.1093/nar/19.21.6059. PMC 329079. PMID 1682898.
- Williamson P, Lang J, Boyd Y (November 1991). "The gonadotropin-releasing hormone (Gnrh) gene maps to mouse chromosome 14 and identifies a homologous region on human chromosome 8". Somatic Cell and Molecular Genetics. 17 (6): 609–15. doi:10.1007/BF01233626. PMID 1767338.
- Hayflick JS, Adelman JP, Seeburg PH (August 1989). "The complete nucleotide sequence of the human gonadotropin-releasing hormone gene". Nucleic Acids Research. 17 (15): 6403–4. doi:10.1093/nar/17.15.6403. PMC 318303. PMID 2671939.
- Nikolics K, Mason AJ, Szönyi E, Ramachandran J, Seeburg PH (1985). "A prolactin-inhibiting factor within the precursor for human gonadotropin-releasing hormone". Nature. 316 (6028): 511–7. doi:10.1038/316511a0. PMID 2863757.
- Adelman JP, Mason AJ, Hayflick JS, Seeburg PH (January 1986). "Isolation of the gene and hypothalamic cDNA for the common precursor of gonadotropin-releasing hormone and prolactin release-inhibiting factor in human and rat". Proceedings of the National Academy of Sciences of the United States of America. 83 (1): 179–83. doi:10.1073/pnas.83.1.179. PMC 322815. PMID 2867548.
- Yang-Feng TL, Seeburg PH, Francke U (January 1986). "Human luteinizing hormone-releasing hormone gene (LHRH) is located on short arm of chromosome 8 (region 8p11.2----p21)". Somatic Cell and Molecular Genetics. 12 (1): 95–100. doi:10.1007/BF01560732. PMID 3511544.
- Seeburg PH, Adelman JP (1984). "Characterization of cDNA for precursor of human luteinizing hormone releasing hormone". Nature. 311 (5987): 666–8. doi:10.1038/311666a0. PMID 6090951.
- Tan L, Rousseau P (December 1982). "The chemical identity of the immunoreactive LHRH-like peptide biosynthesized in the human placenta". Biochemical and Biophysical Research Communications. 109 (3): 1061–71. doi:10.1016/0006-291X(82)92047-2. PMID 6760865.
- Dong KW, Yu KL, Roberts JL (December 1993). "Identification of a major up-stream transcription start site for the human progonadotropin-releasing hormone gene used in reproductive tissues and cell lines". Molecular Endocrinology. 7 (12): 1654–66. doi:10.1210/me.7.12.1654. PMID 8145771.
- Kakar SS, Jennes L (November 1995). "Expression of gonadotropin-releasing hormone and gonadotropin-releasing hormone receptor mRNAs in various non-reproductive human tissues". Cancer Letters. 98 (1): 57–62. doi:10.1016/S0304-3835(06)80010-8. PMID 8529206.
- Nagy A, Schally AV, Armatis P, Szepeshazi K, Halmos G, Kovacs M, Zarandi M, Groot K, Miyazaki M, Jungwirth A, Horvath J (July 1996). "Cytotoxic analogs of luteinizing hormone-releasing hormone containing doxorubicin or 2-pyrrolinodoxorubicin, a derivative 500-1000 times more potent". Proceedings of the National Academy of Sciences of the United States of America. 93 (14): 7269–73. doi:10.1073/pnas.93.14.7269. PMC 38972. PMID 8692981.
- Chegini N, Rong H, Dou Q, Kipersztok S, Williams RS (September 1996). "Gonadotropin-releasing hormone (GnRH) and GnRH receptor gene expression in human myometrium and leiomyomata and the direct action of GnRH analogs on myometrial smooth muscle cells and interaction with ovarian steroids in vitro". The Journal of Clinical Endocrinology and Metabolism. 81 (9): 3215–21. doi:10.1210/jc.81.9.3215. PMID 8784072.
- Bonaldo MF, Lennon G, Soares MB (September 1996). "Normalization and subtraction: two approaches to facilitate gene discovery". Genome Research. 6 (9): 791–806. doi:10.1101/gr.6.9.791. PMID 8889548.
- Dong KW, Yu KL, Chen ZG, Chen YD, Roberts JL (July 1997). "Characterization of multiple promoters directing tissue-specific expression of the human gonadotropin-releasing hormone gene". Endocrinology. 138 (7): 2754–62. doi:10.1210/en.138.7.2754. PMID 9202214.
- Twan WH, Hwang JS, Lee YH, Jeng SR, Yueh WS, Tung YH, Wu HF, Dufour S, Chang CF (January 2006). "The presence and ancestral role of gonadotropin-releasing hormone in the reproduction of scleractinian coral, Euphyllia ancora". Endocrinology. 147 (1): 397–406. doi:10.1210/en.2005-0584. PMID 16195400.
External links
- Overview of all the structural information available in the PDB for UniProt: P01148 (Progonadoliberin-1) at the PDBe-KB.