Globin
The globins are a superfamily of heme-containing globular proteins, involved in binding and/or transporting oxygen. These proteins all incorporate the globin fold, a series of eight alpha helical segments. Two prominent members include myoglobin and hemoglobin. Both of these proteins reversibly bind oxygen via a heme prosthetic group. They are widely distributed in many organisms.[2]
| Globin family | |||||||||
|---|---|---|---|---|---|---|---|---|---|
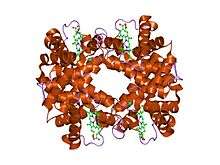 the Structure of deoxyhemoglobin Rothschild 37 beta Trp----Arg: a mutation that creates an intersubunit chloride-binding site.[1] | |||||||||
| Identifiers | |||||||||
| Symbol | Globin | ||||||||
| Pfam | PF00042 | ||||||||
| Pfam clan | CL0090 | ||||||||
| InterPro | IPR000971 | ||||||||
| PROSITE | PS01033 | ||||||||
| SCOPe | 1hba / SUPFAM | ||||||||
| CDD | cd01067 | ||||||||
| |||||||||
| Bacterial-like Globin | |||||||||
|---|---|---|---|---|---|---|---|---|---|
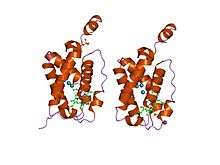 crystal structure of "truncated" hemoglobin n (hbn) from mycobacterium tuberculosis, soaked with xe atoms | |||||||||
| Identifiers | |||||||||
| Symbol | Bac_globin | ||||||||
| Pfam | PF01152 | ||||||||
| Pfam clan | CL0090 | ||||||||
| InterPro | IPR001486 | ||||||||
| PROSITE | PDOC00933 | ||||||||
| SCOPe | 1dlw / SUPFAM | ||||||||
| |||||||||
Structure
Globin superfamily members share a common three-dimensional fold.[3] This 'globin fold' typically consists of eight alpha helices, although some proteins have additional helix extensions at their termini.[4] Since the globin fold contains only helices, it is classified as an all-alpha protein fold.
The globin fold is found in its namesake globin families as well as in phycocyanins. The globin fold was thus the first protein fold discovered (myoglobin was the first protein whose structure was solved).
Helix packing
The eight helices of the globin fold core share significant nonlocal structure, unlike other structural motifs in which amino acids close to each other in primary sequence are also close in space. The helices pack together at an average angle of about 50 degrees, significantly steeper than other helical packings such as the helix bundle. The exact angle of helix packing depends on the sequence of the protein, because packing is mediated by the sterics and hydrophobic interactions of the amino acid side chains near the helix interfaces.
Evolution
Globins evolved from a common ancestor and can be divided into three groups: single-domain globins, and two types of chimeric globins, flavohaemoglobins and globin-coupled sensors. Bacteria have all three types of globins, while archaea lack flavohaemoglobins, and eukaryotes lack globin-coupled sensors.[5] Several functionally different haemoglobins can coexist in the same species.
Eight globins are known to occur in vertebrates: androglobin, cytoglobin, globin E, globin X, globin Y, hemoglobin, myoglobin and neuroglobin.
Sequence conservation
Although the fold of the globin superfamily is highly evolutionarily conserved, the sequences that form the fold can have as low as 16% sequence identity. While the sequence specificity of the fold is not stringent, the hydrophobic core of the protein must be maintained and hydrophobic patches on the generally hydrophilic solvent-exposed surface must be avoided in order for the structure to remain stable and soluble. The most famous mutation in the globin fold is a change from glutamate to valine in one chain of the hemoglobin molecule. This mutation creates a "hydrophobic patch" on the protein surface that promotes intermolecular aggregation, the molecular event that gives rise to sickle-cell anemia.
Subfamilies
- Leghaemoglobin InterPro: IPR001032
- Myoglobin InterPro: IPR002335
- Erythrocruorin InterPro: IPR002336
- Hemoglobin, beta InterPro: IPR002337
- Hemoglobin, alpha InterPro: IPR002338
- Myoglobin, trematode type InterPro: IPR011406
- Globin, nematode InterPro: IPR012085
- Globin, lamprey/hagfish type InterPro: IPR013314
- Globin, annelid-type InterPro: IPR013316
- Haemoglobin, extracellular InterPro: IPR014610
Examples
Human genes encoding globin proteins include:
The globins include:
- Haemoglobin (Hb)
- Myoglobin (Mb)
- Neuroglobin: a myoglobin-like haemprotein expressed in vertebrate brain and retina, where it is involved in neuroprotection from damage due to hypoxia or ischemia.[6] Neuroglobin belongs to a branch of the globin family that diverged early in evolution.
- Cytoglobin: an oxygen sensor expressed in multiple tissues. Related to neuroglobin.[7]
- Erythrocruorin: highly cooperative extracellular respiratory proteins found in annelids and arthropods that are assembled from as many as 180 subunit into hexagonal bilayers.[8]
- Leghaemoglobin (legHb or symbiotic Hb): occurs in the root nodules of leguminous plants, where it facilitates the diffusion of oxygen to symbiotic bacteriods in order to promote nitrogen fixation.
- Non-symbiotic haemoglobin (NsHb): occurs in non-leguminous plants, and can be over-expressed in stressed plants .
- Flavohaemoglobins (FHb): chimeric, with an N-terminal globin domain and a C-terminal ferredoxin reductase-like NAD/FAD-binding domain. FHb provides protection against nitric oxide via its C-terminal domain, which transfers electrons to haem in the globin.[9]
- Globin E: a globin responsible for storing and delivering oxygen to the retina in birds[10]
- Globin-coupled sensors: chimeric, with an N-terminal myoglobin-like domain and a C-terminal domain that resembles the cytoplasmic signalling domain of bacterial chemoreceptors. They bind oxygen, and act to initiate an aerotactic response or regulate gene expression.[11][12]
- Protoglobin: a single domain globin found in archaea that is related to the N-terminal domain of globin-coupled sensors.[13]
- Truncated 2/2 globin: lack the first helix, giving them a 2-over-2 instead of the canonical 3-over-3 alpha-helical sandwich fold. Can be divided into three main groups (I, II and II) based on structural features.
- HbN (or GlbN): a truncated haemoglobin-like protein that binds oxygen cooperatively with a very high affinity and a slow dissociation rate, which may exclude it from oxygen transport. It appears to be involved in bacterial nitric oxide detoxification and in nitrosative stress.[14]
- Cyanoglobin (or GlbN): a truncated haemoprotein found in cyanobacteria that has high oxygen affinity, and which appears to serve as part of a terminal oxidase, rather than as a respiratory pigment.[15]
- HbO (or GlbO): a truncated haemoglobin-like protein with a lower oxygen affinity than HbN. HbO associates with the bacterial cell membrane, where it significantly increases oxygen uptake over membranes lacking this protein. HbO appears to interact with a terminal oxidase, and could participate in an oxygen/electron-transfer process that facilitates oxygen transfer during aerobic metabolism.[16]
- Glb3: a nuclear-encoded truncated haemoglobin from plants that appears more closely related to HbO than HbN. Glb3 from Arabidopsis thaliana (Mouse-ear cress) exhibits an unusual concentration-independent binding of oxygen and carbon dioxide.[17]
References
- Kavanaugh JS, Rogers PH, Case DA, Arnone A (April 1992). "High-resolution X-ray study of deoxyhemoglobin Rothschild 37 beta Trp----Arg: a mutation that creates an intersubunit chloride-binding site". Biochemistry. 31 (16): 4111–21. doi:10.1021/bi00131a030. PMID 1567857.
- Vinogradov SN, Hoogewijs D, Bailly X, Mizuguchi K, Dewilde S, Moens L, Vanfleteren JR (August 2007). "A model of globin evolution". Gene. 398 (1–2): 132–42. doi:10.1016/j.gene.2007.02.041. PMID 17540514.
- Branden, Carl; Tooze, John (1999). Introduction to protein structure (2nd ed.). New York: Garland Pub. ISBN 978-0815323051.
- Bolognesi, M; Onesti, S; Gatti, G; Coda, A; Ascenzi, P; Brunori, M (1989). "Aplysia limacina myoglobin. Crystallographic analysis at 1.6 a resolution". Journal of Molecular Biology. 205 (3): 529–44. doi:10.1016/0022-2836(89)90224-6. PMID 2926816.
- Vinogradov SN, Hoogewijs D, Bailly X, Arredondo-Peter R, Gough J, Dewilde S, Moens L, Vanfleteren JR (2006). "A phylogenomic profile of globins". BMC Evol. Biol. 6: 31. doi:10.1186/1471-2148-6-31. PMC 1457004. PMID 16600051.
- Pesce A, Dewilde S, Nardini M, Moens L, Ascenzi P, Hankeln T, Burmester T, Bolognesi M (September 2003). "Human brain neuroglobin structure reveals a distinct mode of controlling oxygen affinity". Structure. 11 (9): 1087–95. doi:10.1016/S0969-2126(03)00166-7. PMID 12962627.
- Fago A, Hundahl C, Malte H, Weber RE (2004). "Functional properties of neuroglobin and cytoglobin. Insights into the ancestral physiological roles of globins". IUBMB Life. 56 (11–12): 689–96. doi:10.1080/15216540500037299. PMID 15804833.
- Royer WE, Omartian MN, Knapp JE (January 2007). "Low resolution crystal structure of Arenicola erythrocruorin: influence of coiled coils on the architecture of a megadalton respiratory protein". J. Mol. Biol. 365 (1): 226–36. doi:10.1016/j.jmb.2006.10.016. PMC 1847385. PMID 17084861.
- Mukai M, Mills CE, Poole RK, Yeh SR (March 2001). "Flavohemoglobin, a globin with a peroxidase-like catalytic site". J. Biol. Chem. 276 (10): 7272–7. doi:10.1074/jbc.M009280200. PMID 11092893.
- Blank M, Kiger L, Thielebein A, Gerlach F, Hankeln T, Marden MC, Burmeister T (2011). "Oxygen supply from the bird's eye perspective: Globin E is a respiratory protein in the chicken retina". J. Biol. Chem. 286 (30): 26507–15. doi:10.1074/jbc.M111.224634. PMC 3143615. PMID 21622558.
- Hou S, Freitas T, Larsen RW, Piatibratov M, Sivozhelezov V, Yamamoto A, Meleshkevitch EA, Zimmer M, Ordal GW, Alam M (July 2001). "Globin-coupled sensors: a class of heme-containing sensors in Archaea and Bacteria". Proc. Natl. Acad. Sci. U.S.A. 98 (16): 9353–8. doi:10.1073/pnas.161185598. PMC 55424. PMID 11481493.
- Freitas TA, Saito JA, Hou S, Alam M (January 2005). "Globin-coupled sensors, protoglobins, and the last universal common ancestor". J. Inorg. Biochem. 99 (1): 23–33. doi:10.1016/j.jinorgbio.2004.10.024. PMID 15598488.
- Freitas TA, Hou S, Dioum EM, Saito JA, Newhouse J, Gonzalez G, Gilles-Gonzalez MA, Alam M (April 2004). "Ancestral hemoglobins in Archaea". Proc. Natl. Acad. Sci. U.S.A. 101 (17): 6675–80. doi:10.1073/pnas.0308657101. PMC 404104. PMID 15096613.
- Lama A, Pawaria S, Dikshit KL (July 2006). "Oxygen binding and NO scavenging properties of truncated hemoglobin, HbN, of Mycobacterium smegmatis". FEBS Lett. 580 (17): 4031–41. doi:10.1016/j.febslet.2006.06.037. PMID 16814781.
- Yeh DC, Thorsteinsson MV, Bevan DR, Potts M, La Mar GN (February 2000). "Solution 1H NMR study of the heme cavity and folding topology of the abbreviated chain 118-residue globin from the cyanobacterium Nostoc commune". Biochemistry. 39 (6): 1389–99. doi:10.1021/bi992081l. PMID 10684619.
- Pathania R, Navani NK, Rajamohan G, Dikshit KL (May 2002). "Mycobacterium tuberculosis hemoglobin HbO associates with membranes and stimulates cellular respiration of recombinant Escherichia coli". J. Biol. Chem. 277 (18): 15293–302. doi:10.1074/jbc.M111478200. PMID 11796724.
- Watts RA, Hunt PW, Hvitved AN, Hargrove MS, Peacock WJ, Dennis ES (August 2001). "A hemoglobin from plants homologous to truncated hemoglobins of microorganisms". Proc. Natl. Acad. Sci. U.S.A. 98 (18): 10119–24. doi:10.1073/pnas.191349198. PMC 56925. PMID 11526234.