Ploidy
Ploidy (/ˈplɔɪdi/) is the number of complete sets of chromosomes in a cell, and hence the number of possible alleles for autosomal and pseudoautosomal genes. Somatic cells, tissues, and individual organisms can be described according to the number of sets of chromosomes present (the "ploidy level"): monoploid (1 set), diploid (2 sets), triploid (3 sets), tetraploid (4 sets), pentaploid (5 sets), hexaploid (6 sets), heptaploid[2] or septaploid[3] (7 sets), etc. The generic term polyploid is often used to describe cells with three or more chromosome sets.[4][5]

Virtually all sexually reproducing organisms are made up of somatic cells that are diploid or greater, but ploidy level may vary widely between different organisms, between different tissues within the same organism, and at different stages in an organism's life cycle. Half of all known plant genera contain polyploid species, and about two-thirds of all grasses are polyploid.[6] Many animals are uniformly diploid, though polyploidy is common in invertebrates, reptiles, and amphibians. In some species, ploidy varies between individuals of the same species (as in the social insects), and in others entire tissues and organ systems may be polyploid despite the rest of the body being diploid (as in the mammalian liver). For many organisms, especially plants and fungi, changes in ploidy level between generations are major drivers of speciation. In mammals and birds, ploidy changes are typically fatal.[7] There is, however, evidence of polyploidy in organisms now considered to be diploid, suggesting that polyploidy has contributed to evolutionary diversification in plants and animals through successive rounds of polyploidization and rediploidization.[8][9]
Humans are diploid organisms, carrying two complete sets of chromosomes in their somatic cells: one set of 23 chromosomes from their father and one set of 23 chromosomes from their mother. The two sets combined provide a full complement of 46 chromosomes. This total number of individual chromosomes (counting all complete sets) is called the chromosome number. The number of chromosomes found in a single complete set of chromosomes is called the monoploid number (x). The haploid number (n) refers to the total number of chromosomes found in a gamete (a sperm or egg cell produced by meiosis in preparation for sexual reproduction). Under normal conditions, the haploid number is exactly half the total number of chromosomes present in the organism's somatic cells. For diploid organisms, the monoploid number and haploid number are equal; in humans, both are equal to 23. When a human germ cell undergoes meiosis, the diploid 46-chromosome complement is split in half to form haploid gametes. After fusion of a male and a female gamete (each containing 1 set of 23 chromosomes) during fertilization, the resulting zygote again has the full complement of 46 chromosomes: 2 sets of 23 chromosomes.
Etymology
The term ploidy is a back-formation from haploidy and diploidy. "Ploid" is a combination of Ancient Greek -πλόος (-plóos, “-fold”) and -ειδής (-eidḗs), from εἶδος (eîdos, "form, likeness").[lower-alpha 1] The principal meaning of the Greek word ᾰ̔πλόος (haplóos) is "single",[10] from ἁ- (ha-, “one, same”).[11] διπλόος (diplóos) means "duplex" or "two-fold". Diploid therefore means "duplex-shaped" (compare "humanoid", "human-shaped").
Polish botanist Eduard Strasburger coined the terms haploid and diploid in 1905.[lower-alpha 2] Some authors suggest that Strasburger based the terms on August Weismann's conception of the id (or germ plasm),[14][15][16] hence haplo-id and diplo-id. The two terms were brought into the English language from German through William Henry Lang's 1908 translation of a 1906 textbook by Strasburger and colleagues.[17]
Types of ploidy
Haploid and monoploid
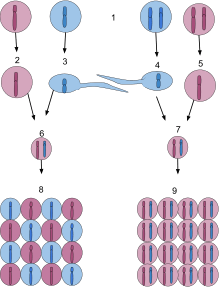
1) A haploid organism is on the left and a diploid organism is on the right.
2 and 3) Haploid egg and sperm carrying the dominant purple gene and the recessive blue gene, respectively. These gametes are produced by simple mitosis of cells in the germ line.
4 and 5) Diploid sperm and egg carrying the recessive blue gene and the dominant purple gene, respectively. These gametes are produced by meiosis, which halves the number of chromosomes in the diploid germ cells.
6) The short-lived diploid state of haploid organisms, a zygote generated by the union of two haploid gametes during sex.
7) The diploid zygote which has just been fertilized by the union of haploid egg and sperm during sex.
8) Cells of the diploid structure quickly undergo meiosis to produce spores containing the meiotically halved number of chromosomes, restoring haploidy. These spores express either the mother's dominant gene or the father's recessive gene and proceed by mitotic division to build a new entirely haploid organism.
9) The diploid zygote proceeds by mitotic division to build a new entirely diploid organism. These cells possess both the purple and blue genes, but only the purple gene is expressed since it is dominant over the recessive blue gene.
The term haploid is used with two distinct but related definitions. In the most generic sense, haploid refers to having the number of sets of chromosomes normally found in a gamete.[18] Because two gametes necessarily combine during sexual reproduction to form a single zygote from which somatic cells are generated, healthy gametes always possess exactly half the number of sets of chromosomes found in the somatic cells, and therefore "haploid" in this sense refers to having exactly half the number of sets of chromosomes found in a somatic cell. By this definition, an organism whose gametic cells contain a single copy of each chromosome (one set of chromosomes) may be considered haploid while the somatic cells, containing two copies of each chromosome (two sets of chromosomes), are diploid. This scheme of diploid somatic cells and haploid gametes is widely used in the animal kingdom and is the simplest to illustrate in diagrams of genetics concepts. But this definition also allows for haploid gametes with more than one set of chromosomes. As given above, gametes are by definition haploid, regardless of the actual number of sets of chromosomes they contain. An organism whose somatic cells are tetraploid (four sets of chromosomes), for example, will produce gametes by meiosis that contain two sets of chromosomes. These gametes might still be called haploid even though they are numerically diploid.
An alternative usage defines "haploid" as having a single copy of each chromosome – that is, one and only one set of chromosomes.[19] In this case, the nucleus of a eukaryotic cell is only said to be haploid if it has a single set of chromosomes, each one not being part of a pair. By extension a cell may be called haploid if its nucleus has one set of chromosomes, and an organism may be called haploid if its body cells (somatic cells) have one set of chromosomes per cell. By this definition haploid therefore would not be used to refer to the gametes produced by the tetraploid organism in the example above, since these gametes are numerically diploid. The term monoploid is often used as a less ambiguous way to describe a single set of chromosomes; by this second definition, haploid and monoploid are identical and can be used interchangeably.
Gametes (sperm and ova) are haploid cells. The haploid gametes produced by most organisms combine to form a zygote with n pairs of chromosomes, i.e. 2n chromosomes in total. The chromosomes in each pair, one of which comes from the sperm and one from the egg, are said to be homologous. Cells and organisms with pairs of homologous chromosomes are called diploid. For example, most animals are diploid and produce haploid gametes. During meiosis, sex cell precursors have their number of chromosomes halved by randomly "choosing" one member of each pair of chromosomes, resulting in haploid gametes. Because homologous chromosomes usually differ genetically, gametes usually differ genetically from one another.
All plants and many fungi and algae switch between a haploid and a diploid state, with one of the stages emphasized over the other. This is called alternation of generations. Most fungi and algae are haploid during the principal stage of their life cycle, as are some primitive plants like mosses. More recently evolved plants, like the gymnosperms and angiosperms, spend the majority of their life cycle in the diploid stage. Most animals are diploid, but male bees, wasps, and ants are haploid organisms because they develop from unfertilized, haploid eggs, while females (workers and queens) are diploid, making their system haplodiploid.
In some cases there is evidence that the n chromosomes in a haploid set have resulted from duplications of an originally smaller set of chromosomes. This "base" number – the number of apparently originally unique chromosomes in a haploid set – is called the monoploid number,[20] also known as basic or cardinal number,[21] or fundamental number.[22][23] As an example, the chromosomes of common wheat are believed to be derived from three different ancestral species, each of which had 7 chromosomes in its haploid gametes. The monoploid number is thus 7 and the haploid number is 3 × 7 = 21. In general n is a multiple of x. The somatic cells in a wheat plant have six sets of 7 chromosomes: three sets from the egg and three sets from the sperm which fused to form the plant, giving a total of 42 chromosomes. As a formula, for wheat 2n = 6x = 42, so that the haploid number n is 21 and the monoploid number x is 7. The gametes of common wheat are considered to be haploid, since they contain half the genetic information of somatic cells, but they are not monoploid, as they still contain three complete sets of chromosomes (n = 3x).[24]
In the case of wheat, the origin of its haploid number of 21 chromosomes from three sets of 7 chromosomes can be demonstrated. In many other organisms, although the number of chromosomes may have originated in this way, this is no longer clear, and the monoploid number is regarded as the same as the haploid number. Thus in humans, x = n = 23.
Diploid
Diploid cells have two homologous copies of each chromosome, usually one from the mother and one from the father. All or nearly all mammals are diploid organisms. The suspected tetraploid (possessing four chromosome sets) plains viscacha rat (Tympanoctomys barrerae) and golden viscacha rat (Pipanacoctomys aureus)[25] have been regarded as the only known exceptions (as of 2004).[26] However, some genetic studies have rejected any polyploidism in mammals as unlikely, and suggest that amplification and dispersion of repetitive sequences best explain the large genome size of these two rodents.[27] All normal diploid individuals have some small fraction of cells that display polyploidy. Human diploid cells have 46 chromosomes (the somatic number, 2n) and human haploid gametes (egg and sperm) have 23 chromosomes (n). Retroviruses that contain two copies of their RNA genome in each viral particle are also said to be diploid. Examples include human foamy virus, human T-lymphotropic virus, and HIV.[28]
Polyploidy
Polyploidy is the state where all cells have multiple sets of chromosomes beyond the basic set, usually 3 or more. Specific terms are triploid (3 sets), tetraploid (4 sets), pentaploid (5 sets), hexaploid (6 sets), heptaploid[2] or septaploid[3] (7 sets), octoploid (8 sets), nonaploid (9 sets), decaploid (10 sets), undecaploid (11 sets), dodecaploid (12 sets), tridecaploid (13 sets), tetradecaploid (14 sets), etc.[29][30][31][32] Some higher ploidies include hexadecaploid (16 sets), dotriacontaploid (32 sets), and tetrahexacontaploid (64 sets),[33] though Greek terminology may be set aside for readability in cases of higher ploidy (such as "16-ploid").[31] Polytene chromosomes of plants and fruit flies can be 1024-ploid.[34][35] Ploidy of systems such as the salivary gland, elaiosome, endosperm, and trophoblast can exceed this, up to 1048576-ploid in the silk glands of the commercial silkworm Bombyx mori.[36]
The chromosome sets may be from the same species or from closely related species. In the latter case, these are known as allopolyploids (or amphidiploids, which are allopolyploids that behave as if they were normal diploids). Allopolyploids are formed from the hybridization of two separate species. In plants, this probably most often occurs from the pairing of meiotically unreduced gametes, and not by diploid–diploid hybridization followed by chromosome doubling.[37] The so-called Brassica triangle is an example of allopolyploidy, where three different parent species have hybridized in all possible pair combinations to produce three new species.
Polyploidy occurs commonly in plants, but rarely in animals. Even in diploid organisms, many somatic cells are polyploid due to a process called endoreduplication, where duplication of the genome occurs without mitosis (cell division). The extreme in polyploidy occurs in the fern genus Ophioglossum, the adder's-tongues, in which polyploidy results in chromosome counts in the hundreds, or, in at least one case, well over one thousand.
It is possible for polyploid organisms to revert to lower ploidy by haploidisation.
In bacteria and archaea
Polyploidy is a characteristic of the bacterium Deinococcus radiodurans [38] and of the archaeon Halobacterium salinarum.[39] These two species are highly resistant to ionizing radiation and desiccation, conditions that induce DNA double-strand breaks.[40][41] This resistance appears to be due to efficient homologous recombinational repair.
Variable or indefinite ploidy
Depending on growth conditions, prokaryotes such as bacteria may have a chromosome copy number of 1 to 4, and that number is commonly fractional, counting portions of the chromosome partly replicated at a given time. This is because under exponential growth conditions the cells are able to replicate their DNA faster than they can divide.
In ciliates, the macronucleus is called ampliploid, because only part of the genome is amplified.[42]
Mixoploidy
Mixoploidy is the case where two cell lines, one diploid and one polyploid, coexist within the same organism. Though polyploidy in humans is not viable, mixoploidy has been found in live adults and children.[43] There are two types: diploid-triploid mixoploidy, in which some cells have 46 chromosomes and some have 69,[44] and diploid-tetraploid mixoploidy, in which some cells have 46 and some have 92 chromosomes. It is a major topic of cytology.
Dihaploidy and polyhaploidy
Dihaploid and polyhaploid cells are formed by haploidisation of polyploids, i.e., by halving the chromosome constitution.
Dihaploids (which are diploid) are important for selective breeding of tetraploid crop plants (notably potatoes), because selection is faster with diploids than with tetraploids. Tetraploids can be reconstituted from the diploids, for example by somatic fusion.
The term "dihaploid" was coined by Bender[45] to combine in one word the number of genome copies (diploid) and their origin (haploid). The term is well established in this original sense,[46][47] but it has also been used for doubled monoploids or doubled haploids, which are homozygous and used for genetic research.[48]
Euploidy and aneuploidy
Euploidy (Greek eu, "true" or "even") is the state of a cell or organism having one or more than one set of the same set of chromosomes, possibly excluding the sex-determining chromosomes. For example, most human cells have 2 of each of the 23 homologous monoploid chromosomes, for a total of 46 chromosomes. A human cell with one extra set of the 23 normal chromosomes (functionally triploid) would be considered euploid. Euploid karyotypes would consequentially be a multiple of the haploid number, which in humans is 23.
Aneuploidy is the state where one or more individual chromosomes of a normal set are absent or present in more than their usual number of copies (excluding the absence or presence of complete sets, which is considered euploidy). Unlike euploidy, aneuploid karyotypes will not be a multiple of the haploid number. In humans, examples of aneuploidy include having a single extra chromosome (as in Down syndrome, where affected individuals have three copies of chromosome 21) or missing a chromosome (as in Turner syndrome, where affected individuals are missing an X chromosome). Aneuploid karyotypes are given names with the suffix -somy (rather than -ploidy, used for euploid karyotypes), such as trisomy and monosomy.
Homoploid
Homoploid means "at the same ploidy level", i.e. having the same number of homologous chromosomes. For example, homoploid hybridization is hybridization where the offspring have the same ploidy level as the two parental species. This contrasts with a common situation in plants where chromosome doubling accompanies or occurs soon after hybridization. Similarly, homoploid speciation contrasts with polyploid speciation.
Zygoidy and azygoidy
Zygoidy is the state in which the chromosomes are paired and can undergo meiosis. The zygoid state of a species may be diploid or polyploid.[49][50] In the azygoid state the chromosomes are unpaired. It may be the natural state of some asexual species or may occur after meiosis. In diploid organisms the azygoid state is monoploid. (See below for dihaploidy.)
Special cases
More than one nucleus per cell
In the strictest sense, ploidy refers to the number of sets of chromosomes in a single nucleus rather than in the cell as a whole. Because in most situations there is only one nucleus per cell, it is commonplace to speak of the ploidy of a cell, but in cases in which there is more than one nucleus per cell, more specific definitions are required when ploidy is discussed. Authors may at times report the total combined ploidy of all nuclei present within the cell membrane of a syncytium,[36] though usually the ploidy of each nucleus is described individually. For example, a fungal dikaryon with two separate haploid nuclei is distinguished from a diploid cell in which the chromosomes share a nucleus and can be shuffled together.[51]
Ancestral ploidy levels
It is possible on rare occasions for ploidy to increase in the germline, which can result in polyploid offspring and ultimately polyploid species. This is an important evolutionary mechanism in both plants and animals and is known as a primary driver of speciation.[8] As a result, it may become desirable to distinguish between the ploidy of a species or variety as it presently breeds and that of an ancestor. The number of chromosomes in the ancestral (non-homologous) set is called the monoploid number (x), and is distinct from the haploid number (n) in the organism as it now reproduces.
Common wheat (Triticum aestivum) is an organism in which x and n differ. Each plant has a total of six sets of chromosomes (with two sets likely having been obtained from each of three different diploid species that are its distant ancestors). The somatic cells are hexaploid, 2n = 6x = 42 (where the monoploid number x = 7 and the haploid number n = 21). The gametes are haploid for their own species, but triploid, with three sets of chromosomes, by comparison to a probable evolutionary ancestor, einkorn wheat.
Tetraploidy (four sets of chromosomes, 2n = 4x) is common in many plant species, and also occurs in amphibians, reptiles, and insects. For example, species of Xenopus (African toads) form a ploidy series, featuring diploid (X. tropicalis, 2n=20), tetraploid (X. laevis, 4n=36), octaploid (X. wittei, 8n=72), and dodecaploid (X. ruwenzoriensis, 12n=108) species.[52]
Over evolutionary time scales in which chromosomal polymorphisms accumulate, these changes become less apparent by karyotype – for example, humans are generally regarded as diploid, but the 2R hypothesis has confirmed two rounds of whole genome duplication in early vertebrate ancestors.
Haplodiploidy
Ploidy can also vary between individuals of the same species or at different stages of the life cycle.[53][54] In some insects it differs by caste. In humans, only the gametes are haploid, but in many of the social insects, including ants, bees, and termites, certain individuals develop from unfertilized eggs, making them haploid for their entire lives, even as adults. In the Australian bulldog ant, Myrmecia pilosula, a haplodiploid species, haploid individuals of this species have a single chromosome and diploid individuals have two chromosomes.[55] In Entamoeba, the ploidy level varies from 4n to 40n in a single population.[56] Alternation of generations occurs in most plants, with individuals "alternating" ploidy level between different stages of their sexual life cycle.
Tissue-specific polyploidy
In large multicellular organisms, variations in ploidy level between different tissues, organs, or cell lineages are common. Because the chromosome number is generally reduced only by the specialized process of meiosis, the somatic cells of the body inherit and maintain the chromosome number of the zygote by mitosis. However, in many situations somatic cells double their copy number by means of endoreduplication as an aspect of cellular differentiation. For example, the hearts of two-year-old human children contain 85% diploid and 15% tetraploid nuclei, but by 12 years of age the proportions become approximately equal, and adults examined contained 27% diploid, 71% tetraploid and 2% octaploid nuclei.[57]
Adaptive and ecological significance of variation in ploidy
There is continued study and debate regarding the fitness advantages or disadvantages conferred by different ploidy levels. A study comparing the karyotypes of endangered or invasive plants with those of their relatives found that being polyploid as opposed to diploid is associated with a 14% lower risk of being endangered, and a 20% greater chance of being invasive.[58] Polyploidy may be associated with increased vigor and adaptability.[59] Some studies suggest that selection is more likely to favor diploidy in host species and haploidy in parasite species.[60]
When a germ cell with an uneven number of chromosomes undergoes meiosis, the chromosomes cannot be evenly divided between the daughter cells, resulting in aneuploid gametes. Triploid organisms, for instance, are usually sterile. Because of this, triploidy is commonly exploited in agriculture to produce seedless fruit such as bananas and watermelons. If the fertilization of human gametes results in three sets of chromosomes, the condition is called triploid syndrome.
Glossary of ploidy numbers
| Term | Description |
|---|---|
| Ploidy number | Number of chromosome sets |
| Monoploid number (x) | Number of chromosomes found in a single complete set |
| Chromosome number | Total number of chromosomes in all sets combined |
| Zygotic number | Number of chromosomes in zygotic cells |
| Haploid or gametic number (n) | Number of chromosomes found in gametes |
| Diploid number | Chromosome number of a diploid organism |
| Tetraploid number | Chromosome number of a tetraploid organism |
The common potato (Solanum tuberosum) is an example of a tetraploid organism, carrying four sets of chromosomes. During sexual reproduction, each potato plant inherits two sets of 12 chromosomes from the pollen parent, and two sets of 12 chromosomes from the ovule parent. The four sets combined provide a full complement of 48 chromosomes. The haploid number (half of 48) is 24. The monoploid number equals the total chromosome number divided by the ploidy level of the somatic cells: 48 chromosomes in total divided by a ploidy level of 4 equals a monoploid number of 12. Hence, the monoploid number (12) and haploid number (24) are distinct in this example.
However, commercial potato crops (as well as many other crop plants) are commonly propagated vegetatively (by asexual reproduction through mitosis),[61] in which case new individuals are produced from a single parent, without the involvement of gametes and fertilization, and all the offspring are genetically identical to each other and to the parent, including in chromosome number. The parents of these vegetative clones may still be capable of producing haploid gametes in preparation for sexual reproduction, but these gametes are not used to create the vegetative offspring by this route.
Specific examples
| Species | Ploidy | Number of chromosomes |
|---|---|---|
| Eucalyptus spp. | Diploid | 2n = 2x = 22 |
| Banana (Musa spp.) | Triploid | 2n = 3x = 33 |
| Coffea arabica | Tetraploid | 2n = 4x = 44 |
| Sequoia sempervirens | Hexaploid | 2n = 6x = 66 |
| Opuntia ficus-indica | Octoploid | 2n = 8x = 88 |
| Species | Number of chromosomes | Ploidy number |
|---|---|---|
| Vinegar/fruit fly | 8 | 2 |
| Wheat | 14, 28 or 42 | 2, 4 or 6 |
| Crocodilian | 32, 34, or 42 | 2 |
| Apple | 34, 51, or 68 | 2, 3 or 4 |
| Human | 46 | 2 |
| Horse | 64 | 2 |
| Chicken | 78 | 2 |
| Gold fish | 100 or more | 2 or polyploid |
Notes
- Compare the etymology of tuple, from the Latin for "-fold".
- The original text in German is as follows: "Schließlich wäre es vielleicht erwünscht, wenn den Bezeichnungen Gametophyt und Sporophyt, die sich allein nur auf Pflanzen mit einfacher und mit doppelter Chromosomenzahl anwenden lassen, solche zur Seite gestellt würden, welche auch für das Tierreich passen. Ich erlaube mir zu diesem Zwecke die Worte Haploid und Diploid, bezw. haploidische und diploidische Generation vorzuschlagen."[12][13]
References
- Daniel Hartl (2011). Essential Genetics: A Genomics Perspective. Jones & Bartlett Learning. p. 177. ISBN 978-0-7637-7364-9.
- U. R. Murty (1973). "Morphology of pachytene chromosomes and its bearing on the nature of polyploidy in the cytological races of Apluda mutica L.". Genetica. 44 (2): 234–243. doi:10.1007/bf00119108.
- Tuguo Tateoka (May 1975). "A contribution to the taxonomy of the Agrostis mertensii-flaccida complex (Poaceae) in Japan". Journal of Plant Research. 88 (2): 65–87. doi:10.1007/bf02491243.
- Rieger, R.; Michaelis, A.; Green, M.M. (1976). Glossary of Genetics and Cytogenetics: Classical and Molecular (4th ed.). Berlin/Heidelberg: Springer-Verlag. p. 434. doi:10.1007/978-3-642-96327-8. ISBN 978-3-540-07668-1.
- Darlington, C. D. (Cyril Dean) (1937). Recent advances in cytology. Philadelphia: P. Blakiston's son & co. p. 60.
- D. Peter Snustad; Michael J. Simmons (2012). Principles of Genetics, 6th edition. John Wiley & Sons. p. 115. ISBN 978-0-470-90359-9.
- Otto, Sarah P. (2007). "The Evolutionary Consequences of Polyploidy". Cell. 131 (3): 452–462. doi:10.1016/j.cell.2007.10.022. ISSN 0092-8674. PMID 17981114.
- Mable, B. K. (2004). "'Why polyploidy is rarer in animals than in plants': myths and mechanisms". Biological Journal of the Linnean Society. 82 (4): 453–466. doi:10.1111/j.1095-8312.2004.00332.x. ISSN 0024-4066.
- Madlung, A (2012). "Polyploidy and its effect on evolutionary success: old questions revisited with new tools". Heredity. 110 (2): 99–104. doi:10.1038/hdy.2012.79. ISSN 0018-067X. PMC 3554449. PMID 23149459.
- "Greek Word Study Tool". www.perseus.tufts.edu.
- "Greek Word Study Tool". www.perseus.tufts.edu.
- Strasburger, Eduard; Allen, Charles E.; Miyake, Kilchi; Overten, James B. (1905). "Histologische Beiträge zur Vererbungsfrage". Jahrbücher für Wissenschaftliche Botanik. 42: 62. Retrieved 2017-03-11.
- Toepfer, Georg (2011). Historisches Worterbüch der Biologie - Geschichte und Theorie der biologischen Grundbegriffe. Stuttgart: J.B. Metzler'sche Verlagsbuchhandlung und Carl Ernst Poeschel Verlag GmbH. p. 169. ISBN 978-3-476-02317-9.
- Battaglia E (2009). "Caryoneme alternative to chromosome and a new caryological nomenclature" (PDF). Caryologia. 62 (4): 48.
- David Haig (2008). "Homologous versus antithetic alternation of generations and the origin of sporophytes" (PDF). The Botanical Review. 74 (3): 395–418. doi:10.1007/s12229-008-9012-x.
- Bennett, Michael D. (2004). "Biological relevance of polyploidy: ecology to genomics". Biological Journal of the Linnean Society. 82 (4): 411–423. doi:10.1111/j.1095-8312.2004.00328.x.
- Strasburger, E.; Noll, F.; Schenck, H.; Karsten, G. 1908. A Textbook of botany, 3rd English ed. (1908) , rev. with the 8th German ed. (1906) , translation by W. H. Lang of Lehrbuch der Botanik für Hochschulen. Macmillan, London.
- "MGI Glossary". Mouse Genome Informatics. Bar Harbor, Maine: The Jackson Laboratory. Retrieved 6 July 2019.
- "Talking Glossary of Genetic Terms". National Human Genome Research Institute. Retrieved 6 July 2019.
- Langlet, 1927.
- Winge, 1917.
- Manton, 1932.
- Fabbri F (1963). "Primo supplemento alle tavole cromosomiche delle Pteridophyta di Alberto Chiarugi". Caryologia. 16: 237–335.
- http://mcb.berkeley.edu/courses/mcb142/lecture%20topics/Amacher/LECTURE_10_CHROM_F08.pdf
- Gallardo MH, González CA, Cebrián I (2006). "Molecular cytogenetics and allotetraploidy in the red vizcacha rat, Tympanoctomys barrerae (Rodentia, Octodontidae)]". Genomics. 88 (2): 214–221. doi:10.1016/j.ygeno.2006.02.010. PMID 16580173.
- Gallardo M. H.; et al. (2004). "Whole-genome duplications in South American desert rodents (Octodontidae)". Biological Journal of the Linnean Society. 82 (4): 443–451. doi:10.1111/j.1095-8312.2004.00331.x.
- Svartman, Marta; Stone, Gary; Stanyon, Roscoe (2005). "Molecular cytogenetics discards polyploidy in mammals". Genomics. 85 (4): 425–430. doi:10.1016/j.ygeno.2004.12.004. PMID 15780745.
- "Archived copy". Archived from the original on 2003-03-30. Retrieved 2008-05-14.CS1 maint: archived copy as title (link)
- Dierschke T, Mandáková T, Lysak MA, Mummenhoff K (September 2009). "A bicontinental origin of polyploid Australian/New Zealand Lepidium species (Brassicaceae)? Evidence from genomic in situ hybridization". Annals of Botany. 104 (4): 681–688. doi:10.1093/aob/mcp161. PMC 2729636. PMID 19589857.
- Simon Renny-Byfield; et al. (2010). "Flow cytometry and GISH reveal mixed ploidy populations and Spartina nonaploids with genomes of S. alterniflora and S. maritima origin". Annals of Botany. 105 (4): 527–533. doi:10.1093/aob/mcq008. PMC 2850792. PMID 20150197.
- Kim E. Hummer; et al. (March 2009). "Decaploidy in Fragaria iturupensis (Rosaceae)". Am. J. Bot. 96 (3): 713–716. doi:10.3732/ajb.0800285. PMID 21628226.
- Talyshinskiĭ, G. M. (1990). "Study of the fractional composition of the proteins in the compound fruit of polyploid mulberry". Shelk (5): 8–10.
- Fujikawa-Yamamoto K (2001). "Temperature dependence in Proliferation of tetraploid Meth-A cells in comparison with the parent diploid cells". Cell Structure and Function. 26 (5): 263–269. doi:10.1247/csf.26.263. PMID 11831358.
- Kiichi Fukui; Shigeki Nakayama (1996). Plant Chromosomes: Laboratory Methods. ISBN 9780849389191.
- "Genes involved in tissue and organ development: Polytene chromosomes, endoreduplication and puffing". The Interactive Fly. Archived from the original on 2005-05-04. Retrieved 2012-12-16.
- Encyclopedia of the Life Sciences (2002) "Polyploidy" Francesco D'Amato and Mauro Durante
- Ramsey, J.; Schemske, D. W. (2002). "Neopolyploidy in Flowering Plants" (PDF). Annual Review of Ecology and Systematics. 33: 589–639. doi:10.1146/annurev.ecolsys.33.010802.150437. Archived from the original (PDF) on 2012-03-19. Retrieved 2015-02-07.
- Hansen MT (1978). "Multiplicity of genome equivalents in the radiation-resistant bacterium Micrococcus radiodurans". J. Bacteriol. 134 (1): 71–75. doi:10.1128/JB.134.1.71-75.1978. PMC 222219. PMID 649572.
- Soppa J (2011). "Ploidy and gene conversion in Archaea". Biochem. Soc. Trans. 39 (1): 150–154. doi:10.1042/BST0390150. PMID 21265763.
- Zahradka K, Slade D, Bailone A, Sommer S, Averbeck D, Petranovic M, Lindner AB, Radman M (2006). "Reassembly of shattered chromosomes in Deinococcus radiodurans". Nature. 443 (7111): 569–573. Bibcode:2006Natur.443..569Z. doi:10.1038/nature05160. PMID 17006450.
- Kottemann M, Kish A, Iloanusi C, Bjork S, DiRuggiero J (2005). "Physiological responses of the halophilic archaeon Halobacterium sp. strain NRC1 to desiccation and gamma irradiation". Extremophiles. 9 (3): 219–227. doi:10.1007/s00792-005-0437-4. PMID 15844015.
- Schaechter, M. Eukaryotic microbes. Amsterdam, Academic Press, 2012, p. 217.
- Edwards MJ; et al. (1994). "Mixoploidy in humans: two surviving cases of diploid-tetraploid mixoploidy and comparison with diploid-triploid mixoploidy". Am J Med Genet. 52 (3): 324–330. doi:10.1002/ajmg.1320520314. PMID 7810564.
- Järvelä, IE; Salo, MK; Santavuori, P; Salonen, RK (1993). "46,XX/69,XXX diploid-triploid mixoploidy with hypothyroidism and precocious puberty". J Med Genet. 30 (11): 966–967. doi:10.1136/jmg.30.11.966. PMC 1016611. PMID 8301657.
- Bender K (1963). "Über die Erzeugung und Entstehung dihaploider Pflanzen bei Solanum tuberosum"". Zeitschrift für Pflanzenzüchtung. 50: 141–166.
- Nogler, G.A. 1984. Gametophytic apomixis. In Embryology of angiosperms. Edited by B.M. Johri. Springer, Berlin, Germany. pp. 475–518.
-
- Pehu E (1996). "The current status of knowledge on the cellular biology of potato". Potato Research. 39 (3): 429–435. doi:10.1007/bf02357948.
-
- Sprague G.F.; Russell W.A.; Penny L.H. (1960). "Mutations affecting quantitative traits in the selfed progeny of double monoploid maize stocks". Genetics. 45 (7): 855–866. PMC 1210096. PMID 17247970.
- Books, Elsevier Science & Technology (1950). Advances in Genetics. Academic Press. ISBN 978-0-12-017603-8.
- Cosín, Darío J. Díaz, Marta Novo, and Rosa Fernández. "Reproduction of Earthworms: Sexual Selection and Parthenogenesis." In Biology of Earthworms, edited by Ayten Karaca, 24:69–86. Berlin, Heidelberg: Springer Berlin Heidelberg, 2011.
- James B. Anderson; Linda M Kohn. "Dikaryons, diploids, and evolution" (PDF). University of Toronto. Archived from the original (PDF) on 2013-05-27. Retrieved 2012-12-16.
- Schmid, M; Evans, BJ; Bogart, JP (2015). "Polyploidy in Amphibia". Cytogenet. Genome Res. 145 (3–4): 315–330. doi:10.1159/000431388. PMID 26112701.
- Parfrey LW, Lahr DJ, Katz LA (2008). "The dynamic nature of eukaryotic genomes". Mol Biol Evol. 25 (4): 787–794. doi:10.1093/molbev/msn032. PMC 2933061. PMID 18258610.
- Qiu Y.-L., Taylor A. B., McManus H. A. (2012). "Evolution of the life cycle in land plants" (PDF). Journal of Systematics and Evolution. 50 (3): 171–194. doi:10.1111/j.1759-6831.2012.00188.x. hdl:2027.42/92043.CS1 maint: multiple names: authors list (link)
- Crosland MW, Crozier RH (1986). "Myrmecia pilosula, an Ant with Only One Pair of Chromosomes". Science. 231 (4743): 1278. Bibcode:1986Sci...231.1278C. doi:10.1126/science.231.4743.1278. PMID 17839565.
- "Archived copy" (PDF). Archived from the original (PDF) on 2014-02-23. Retrieved 2014-02-18.CS1 maint: archived copy as title (link)
- John O. Oberpriller; A Mauro (1991). The Development and Regenerative Potential of Cardiac Muscle. Taylor&Francis. ISBN 9783718605187.
- Pandit, M. K.; Pocock, M. J. O.; Kunin, W. E. (2011-03-28). "Ploidy influences rarity and invasiveness in plants". Journal of Ecology. 99 (5): 1108–1115. doi:10.1111/j.1365-2745.2011.01838.x.
- Gilbert, Natasha (2011-04-06). "Ecologists find genomic clues to invasive and endangered plants". Nature. doi:10.1038/news.2011.213. Retrieved 2011-04-07.
- Nuismer S.; Otto S.P. (2004). "Host-parasite interactions and the evolution of ploidy". Proc. Natl. Acad. Sci. USA. 101 (30): 11036–11039. Bibcode:2004PNAS..10111036N. doi:10.1073/pnas.0403151101. PMC 503737. PMID 15252199.
- "The Biology of Solanum tuberosum (L.) (Potatoes)". Canadian Food Inspection Agency. 2012-03-05.
Sources
- Griffiths, A. J. et al. 2000. An introduction to genetic analysis, 7th ed. W. H. Freeman, New York ISBN 0-7167-3520-2
External links
Some eukaryotic genome-scale or genome size databases and other sources which may list the ploidy levels of many organisms:
- Animal genome size database
- Plant genome size database
- Fungal genome size database
- Protist genome-scale database of Ensembl Genomes
- Nuismer S.; Otto S.P. (2004). "Host-parasite interactions and the evolution of ploidy". Proc. Natl. Acad. Sci. USA. 101 (30): 11036–11039. Bibcode:2004PNAS..10111036N. doi:10.1073/pnas.0403151101. PMC 503737. PMID 15252199. (Supporting Data Set, with information on ploidy level and number of chromosomes of several protists)
- Chromosome number and ploidy mutations YouTube tutorial video