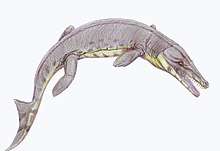
Dyrosauridae
Dyrosauridae is a family of extinct neosuchian crocodyliforms that lived from the Late Cretaceous (Maastrichtian) to the Eocene. Dyrosaurid fossils are globally distributed, having been found in Africa, Asia, Europe, North America and South America. Over a dozen species are currently known, varying greatly in overall size and cranial shape. All were presumably aquatic, with species inhabiting both freshwater and marine environments. Ocean-dwelling dyrosaurids were among the few marine reptiles to survive the Cretaceous–Paleogene extinction event.
| Dyrosauridae | |
|---|---|
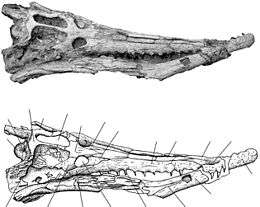 | |
| Skull of the dyrosaurid Arambourgisuchus khouribgaensis | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Suborder: | †Tethysuchia |
| Family: | †Dyrosauridae de Stefano, 1903 |
| Genera | |
The dyrosaurids were a group of mostly marine, long jawed, crocodile-like quadrupeds up to 6 metres (20 ft) long.[1] Based on bone tissue evidence, it has been hypothesized that they were slow-growing[2] near-shore marine animals with interlocking closed jaws,[2] able to swim as well as walk on land. External nostrils at the posterior end of its snout and an internal naris in its pterygoid indicated a habit of hunting while swimming with the top of the head above the water, enabling it to breathe while stalking prey.[1]
Overview

Dyrosaurids are known to have a very characteristic skull shape with a long and thin snout that is approximately 68% of the total skull length. The most anterior part of the skull and snout is the external naris followed posteriorly by two premaxillae bones until they reach two maxillae bones separated by a single nasal bone.[1]
Typical in dyrosaurids is a single nasal element with a characteristic collection of small pits and a constant width until it widens to contact the lacrimal bones, then tapering off for a short distance until it meets the boundary of the frontals and prefrontals.[1]

Dyrosaurs have a premaxilla with shallow pits that extend posterior to the third maxillary alveoli. There are two premaxillae that are narrow as compared to the maxillary bones and extend in the two long maxilla bones which separated by the single nasal bone. The last premaxilla and first maxilla are widely separated by a fourth dentary tooth. Alveoli are widely spaced anteriorly and the space between them decreases posteriorly from the fifteenth alveolus with the diameter remaining constant.[1]
The maxilla is long (approx. two and a half times the length of jugal) and forms most of the lateral margin of the skull.[3] According to Jouve and Barbosa and perhaps depending on the age of the animal, each maxilla bears 13-19 teeth.[3]
An important feature of the dyrosaurid dentition are deep occlusal pits, present particularly in the posterior region of the maxillae that get less pronounced anteriorly. The pits are indicative of an interlocking closed jaw for dyrosaurids because the pits give a way for the upper and lower jaws to alternate. Although they are still present in dyrosaurus phosphaticus, this feature of deep occlusal pits become less developed and broad.[3] Dyrosaurus teeth are homodont, conical, long, and slender, with asymmetrically labial and lingual surfaces. Posterior teeth are shorter and more compressed than anterior tooth indicating that the tooth size decreased anterior to posterior.

Dyrosaurs have a snout to skull length of about 68% and the genus Rhabdognathus and Atlantosuchus, Dyrosaurus and Arambourgisuchus have the largest snout proportions of all dyrosaurids. Snout length was previously used it to establish dyrosaurid relationships, while considering the lengthening of the snout to be a ‘more evolved’ character. This was not congruent with Jouve's conclusion which was that the longest snout is actually the primitive condition so the shorter or longer snout appears independently at least four times in dyrosaurid evolution.[4]
Paleobiogeography
Dyrosaurids were once considered an African group, but discoveries made starting from the 2000s indicate they inhabited the majority of the continents.[5] In fact, basal forms suggest that their cradle may have been North America.
General
| Genus | Status | Age | Location | Description | Images |
|---|---|---|---|---|---|
| Acherontisuchus | Valid | Paleocene | A large-bodied, long-snouted freshwater dyrosaurid from the Cerrejón Formation | ||
| Aigialosuchus | Valid | Campanian | A marine dyrosaurid from the Kristianstad Basin | ||
| Anthracosuchus | Valid | Paleocene | A short-snouted freshwater dyrosaurid from the Cerrejón Formation | ||
| Arambourgisuchus | Valid | Paleocene | A long-snouted marine dyrosaurid | ||
| Atlantosuchus | Valid | Paleocene | A long-snouted marine dyrosaurid with the longest snout length in proportion to body size of any dyrosaurid | ||
| Cerrejonisuchus | Valid | Paleocene | A small-bodied, short-snouted freshwater dyrosaurid from the Cerrejón Formation | ||
| Chenanisuchus | Valid | Maastrichtian-Paleocene | The genus spans the K-Pg boundary |  | |
| Congosaurus | Valid | Paleocene | |||
| Dyrosaurus | Valid | Eocene | A large-bodied, long-snouted marine dyrosaurid | | |
| Fortignathus | Valid | Albian–Cenomanian | A dyrosaurid from the Echkar Formation. | ||
| Guarinisuchus | Valid | Paleocene | A marine dyrosaurid | 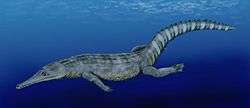 | |
| Hyposaurus | Valid | Maastrichtian-Paleocene | Five species have been named, the most of any dyrosaurid genus; the genus spans the K-Pg boundary | ||
| Phosphatosaurus | Valid | Eocene | A large-bodied, long-snouted marine dyrosaurid with blunt teeth and a spoon-shaped snout tip | ||
| Rhabdognathus | Valid | Maastrichtian-Paleogene | A large-bodied, long-snouted marine dyrosaurid; the genus spans the K-Pg boundary | ||
| Sabinosuchus | Valid | Maastrichtian | A dyrosaurid from the Escondido Formation. | ||
| Sokotosaurus | Junior synonym | Junior synonym of Hyposaurus | |||
| Sokotosuchus | Valid | Maastrichtian | A long-snouted marine dyrosaurid | ||
| Tilemsisuchus | Valid | Eocene | |||
Phylogeny
Jouve et al. (2005) diagnose Dyrosauridae as a clade based on the following seven synapomorphies or shared characters:
- Posteromedial wing of the retroarticular process dorsally situated ventrally on the retroarticular process
- Occipital tuberosities small
- Exoccipital participates largely to the occipital condyle
- Supratemporal fenestra anteroposteriorly strongly elongated
- Symphysis about as wide as high
- Quadratojugal participates largely to the cranial condyle for articulation with the jaw
- 4 premaxillary teeth
Below is a cladogram after Jouve et al. (2005) showing phylogenetic relationships of Dyrosauridae and other closely related neosuchians:
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Composite cladogram for Dyrosauridae (from Jouve et al. 2008 and Barbosa et al. 2008):
| Dyrosauridae |
Dyrosauridae incertae sedis: Tilemsisuchus | |||||||||||||||||||||||||||||||||||||||||||||||||||
Cladogram after Hastings et al. (2011) showing geographic occurrences of taxa:[6]
| Neosuchia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Analysis suggest that the closest relatives of dyrosaurids are Sarcosuchus and Terminonaris.
Palaeobiology
Growth

Evidence for the semi-aquatic life of dyrosaurids comes from careful analysis of bone structure. There are two types of structural bone organization that can occur in aquatic tetrapods: osteoporotic or pachyostotic. Osteoporotic bone is spongy and porous whereas pachyostotic involves an increase in skeletal mass. Spongy/porous bone such as osteoporotic is associated with faster swimming and better maneuverability in water because of the reduction in bone tissue, many extant cetaceans and marine turtles have osteoporotic bone which enables them to be good swimmers. Pachyostotic bone is a general/local increase in skeletal mass which can be caused by osteosclerosis (inner compaction of bone), pachyostosis (hyperplasy of compact cortices) or pachyeosclerosis (combination of the two). Research on dyrosaur bone performed by Rafael César Lima Pedroso de Andrade, Juliana Manso Sayao revealed that this family had osteoporotic bone tissue indicative of a fast-swimming ecology as well as some osteosclerosis which is a component of pachyostotic bone tissue. Osteoporosis is associated with a fully aquatic lifestyle whereas pachyostotic is not fully aquatic but is associated with fast swimming ecology. Therefore, dyrosaurs are semi-aquatic fast swimmers as indicated by their bone structure. Other evidence for near shore, semi-aquatic lifestyle is where the fossils are found, often in transitional marine sediments.[2] -using axial frequency swimming (that used primarily by extant crocodylians) with a greater undulatory motion and frequency of the tail due to highly developed musculature allowing a more powerful forward thrust.[2]
Dyrosaurids have a tissue pattern that is indicative of a slow-growth animal that was determined by careful analysis of a right femur and left tibia. In the left tibia, the cortex had a lamellar zonal bone with five lines of arrested growth (LAGs) which were spaced 300 mm apart, well as a clear vascular networks of primary osteons that decreased in density towards the membrane (periostially). The right femur had double LAGs and an EFS later as well as secondary osteons occurring in the deep cortex and the spongiosa. This tissue growth pattern is a common characteristic of slow growing animals.[2]
Habitat
Dyrosaurids are found in transitional marine sediments from the Late Cretaceous to lower Eocene.[2] This family is known mainly from Maastrichthian deposits in New Jersey and the late Cretaceous to early Paleogene rock from the Tethys Sea in northern and western Africa.[2] Fossils have also been found from the Paleocene and Eocene strata of Pakistan, as well as South America, Brazil, India, Southern Asia as well as coastal. Generally dyrosaurids are recovered from coastal and estuarine deposits through North Africa and the Middle East confirming their existence as semi-aquatic animals.[7]
Dyrosauridae had its greatest taxonomic diversity during the Early Paleogene but it appears as though the clade was able to obtain a greater and more widespread geographic distribution during the Late Cretaceous. The earliest records of dyrosaurids are either in or close to Africa with fragmentary occurrences from the Cenomanian of Sudan and Portugal and several other pre-Maastrichtian, Late Cretaceous discoveries in Egypt. Later, by the Maastrichtian of North America, the record of dyrosaurids became more complete by establishing a widespread distribution that appears to be maintained through the Paleocene and Eocene.[8]
Dyrosaurids have also been found from non-marine sediments. In northern Sudan, dyrosaurids are known from fluvial deposits, indicating that they lived in a river setting.[9] Bones from indeterminate dyrosaurids have been found in inland deposits in Pakistan as well. Some dyrosaurids, such as those from the Umm Himar Formation in Saudi Arabia, inhabited estuarine environments near the coast. The recently named dyrosaurids Cerrejonisuchus and Acherontisuchus have been recovered from the Cerrejón Formation in northwestern Colombia, which is thought to represent a transitional marine-freshwater environment surrounded by rainforest more inland than the estuarine environment of the Umm Himar Formation.[10] Cerrejonisuchus and Acherontisuchus lived in a neotropical setting during a time when global temperatures were much warmer than they are today.[11][12]
Reproduction
In 1978, it was proposed that dyrosaurids lived as adults in the ocean but reproduced in inland freshwater environments. Remains belonging to small-bodied dyrosaurids from Pakistan were interpreted as juveniles. Their presence in inland deposits was viewed as evidence that dyrosaurids hatched far from the ocean.[13] From the lower Eocene Oulad Abdoun Basin, there are very few juvenile dyrosaurids, but numerous similary-sized adult specimens. This has furthered the assumption that juveniles lived in freshwater environments and adults lived in marine environments.[14] Recently however, the large-bodied and fully mature dyrosaurids of the Cerrejón Formation have shown that some dyrosaurids lived their entire lives in inland environments, never returning to the coast.[6]
References
- Jouve, Stephanie. A new description of the skull of Dyrosaurus phosphaticus (Thomas, 1893) (Mesoeucrocodylia: Dyrosauridae) from the Lower Eocene of North Africa. 2005. Retrieved March 2, 2015 from http://cjes.nrc.ca Archived 2002-06-04 at the Wayback Machine.
- Andrade RCLPd, Saya ̃o JM (2014) Paleohistology and Lifestyle Inferences of a Dyrosaurid (Archosauria: Crocodylomorpha) from Paraíba Basin (Northeastern Brazil). PLoS ONE 9(7): e102189. doi:10.1371/journal.pone.0102189
- Antonio Barbosa, José; Wilhelm Armin Kellner, Alexander. Somália Sales Viana, Maria. New dyrosaurid crocodylomorph and evidences for faunal turnover at the K–P transition in Brazil. 2008. Proceedings of the Royal Society. Downloaded from http://rspb.royalsocietypublishing.org/
- Jouve, Stephanie; Bouya, Baadi; Amaghzaz, Mbarek. A LONG-SNOUTED DYROSAURID (CROCODYLIFORMES, MESOEUCROCODYLIA) FROM THE PALEOCENE OF MOROCCO: PHYLOGENETIC AND PALAEOBIOGEOGRAPHIC IMPLICATIONS. 2008. Paleontology, vol. 51, part 2. pp 281-294.
- Jouve et al. (2008)
- Hastings, A.K., Bloch, J. and Jaramillo, C.A. (2011). "A new longirostrine dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleocene of north-eastern Colombia: biogeographic and behavioural implications for new-world dyrosauridae" (PDF). Palaeontology. 54 (5): 1095–116. doi:10.1111/j.1475-4983.2011.01092.x. Archived from the original (PDF) on 31 March 2012. Retrieved 14 Sep 2011.CS1 maint: multiple names: authors list (link)
- Matthew C. Lamanna, Joshua B. Smith, Yousry S. Attia and Peter Dodson Source: Journal of Vertebrate Paleontology, Vol. 24, No. 3 (Sep. 10, 2004), pp. 764-768 Published by: Taylor & Francis, Ltd. on behalf of The Society of Vertebrate Paleontology Stable URL: https://www.jstor.org/stable/4524765 Accessed: 04-03-2015 17:11 UTC.
- Ashu Khosla, Joseph J. W. Sertich, Guntupalli V. R. Prasad and Omkar Verma Source: Journal of Vertebrate Paleontology, Vol. 29, No. 4 (Dec. 12, 2009), pp. 1321-1326 Published by: Taylor & Francis, Ltd. on behalf of The Society of Vertebrate Paleontology Stable URL: https://www.jstor.org/stable/20627142. Accessed: 03-03-2015 07:01 UTC
- Buffetaut, E.; Bussert, R.; Brinkmann, W. (1990). "A new nonmarine vertebrate fauna in the Upper Cretaceous of northern Sudan". Berliner Geowissenschaftlische Abhandlungen. 120: 183–202.
- Hastings, A. K; Bloch, J. I.; Cadena, E. A.; Jaramillo, C. A. (2010). "A new small short-snouted dyrosaurid (Crocodylomorpha, Mesoeucrocodylia) from the Paleocene of northeastern Colombia". Journal of Vertebrate Paleontology. 30 (1): 139–162. doi:10.1080/02724630903409204.
- Head, J. J.; Bloch, J. I.; Hastings, A. K.; Borque, J. R.; Cadena, E. A.; Herrera, F. A.; Polly, P. D.; Jaramillo, C. A. (2009). "Giant boid snake from the Palaeocene neotropics reveals hotter past equatorial temperatures". Nature. 457 (7230): 715–717. Bibcode:2009Natur.457..715H. doi:10.1038/nature07671. PMID 19194448.
- Kanapaux, B. (February 2, 2010). "UF researchers: Ancient crocodile relative likely food source for Titanoboa". University of Florida News. Archived from the original on June 8, 2010. Retrieved February 3, 2010.
- Buffetaut, E. (1978). "Crocodilian remains from the Eocene of Pakistan". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 156: 262–283.
- Jouve, Stephanie; Bardet, Nathalie; Jalil, Nour-Eddine; Superbiola, Xabier Pereda; Bouya, Baâdi and Amagzaz, Mbarek. 2008. The oldest African crocodylian: phylogeny, paleobiogeography, and differential survivorship of marine reptiles through the Cretaceous-Tertiary boundary. Journal of Vertebrate Paleontology 28(2):409–421, June 2008
- Barbosa, J.A., Kellner, A.W.A. and Viana, M.S.S. (2008). New dyrosaurid crocodylomorph and evidences for faunal turnover at the K–P transition in Brazil. Proceedings of the Royal Society B: Biological Sciences: Firstcite
- Buffetaut, E. (1985). L'evolution des crocodiliens. Les animaux disparus-Pour la science, Paris 109.
- Jouve, S.; Bouya, B.; Amaghzaz, M. (2008). "A long-snouted dyrosaurid (Crocodyliformes, Mesoeucrocodylia) from the Paleocene of Morocco: phylogenetic and palaeogeographic implications". Palaeontology. 51 (2): 281–294. doi:10.1111/j.1475-4983.2007.00747.x.
- Jouve, S.; Iarochène, M.; Bouya, B.; Amaghzaz, M. (2005). "A new dyrosaurid crocodyliform from the Palaeocene of Morocco and a phylogenetic analysis of Dyrosauridae". Acta Palaeontologica Polonica. 50 (3): 581–594.
External links
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||