Caytoniales
The Caytoniales (Figs. 1-2) are an extinct order of seed plants known from fossils collected throughout the Mesozoic Era, specifically in the late Triassic to Maastrichtian period, around 250 to 70 million years ago.[2][3][4] They are regarded as seed ferns because they are seed-bearing plants with fern-like leaves.[4] Although at one time considered angiosperms because of their berry-like cupules,[5] that hypothesis was later disproven.[6] Nevertheless, some authorities consider them likely ancestors of angiosperms,[7] whereas others consider angiosperms more likely derived from Glossopteridales.[8] The origin of angiosperms remains an intriguing puzzle (see Evolutionary history of plants).
| Caytoniales | |
|---|---|
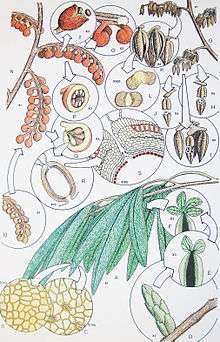 | |
| Fig.1 A whole-plant reconstruction of Caytonia nathorstii[1] | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Division: | †Pteridospermatophyta |
| Order: | †Caytoniales Gothan 1932 |
| Family and form genera | |
| |
History
The first fossils identified in this order were discovered in the Middle Jurassic Gristhorpe bed of the Cloughton formation in Cayton Bay, Yorkshire[9] – hence the name CAYTONiales. They have since been found in Mesozoic rocks all over world.[10] It is likely that Caytoniales flourished in wetland areas, because they are often found with other moisture-loving plants such as horsetails in waterlogged paleosols.[1] The first fossil Caytoniales were preserved as compressions in shale with excellent preservation of cuticles allowing study of cellular histology.[5]
Description
The woody nature of associated stalks and preserved short shoots are evidence that Caytoniales were seasonally deciduous, shrubs and trees.[1] Caytoniales had fertile branches with seed-bearing cupules .[11] The ovules were located inside fleshy cupules with tough outer cuticle. Individual ovules had an apical tube called a micropylar canal, that allowed pollen to pass into the pollen chamber.[12] The outer layers of the cupules were fleshy and fruit-like; it is possible this was to aid in animal dispersal.[1] The cupules are 4-5mm in diameter and about 3 mm long[13] (Fig 1-2),[5] and resemble a blueberry. The extra protection of the reproductive organs gave rise to the idea that Caytoniales were predecessors to angiosperms, which have completely enclosed seeds.
The pollen grains were small, between 25 and 30 μm in diameter. The size of the pollen grains supports the idea that they were wind-pollinated, and their bisaccate wings may have enabled entry into the seed by a pollination drop mechanism.[1] In both respects they were like pollen of pine trees.[4] They were produced in pollen sacs in coalesced groups of four, attached to branching structures. The pollen sacs hang off the structure in clusters, and are typically 2 cm in length.[10]
The most common and widespread part found fossilized are leaves of Sagenopteris (Fig. 3).[5] These are compound leaves consisting of, usually, 4 leaflets arrayed in a palmate manner. The individual leaflets are up to 6 cm in length. The leaflets have anastomosing veins, like those of some ferns, but lacking orders of venation found in angiosperm leaves.[1]
Relationship to Angiosperms
Caytonia was first described by Hamshaw Thomas in 1925. His close examination of the cupules led him to believe this was one of the earliest examples of angiosperms. He mistakenly thought the entire ovule was enclosed in the cupule, unlike typical gymnosperms. He worked meticulously, collecting and cleaning specimens to get the best understanding. He spent weeks boiling fruits in different solutions to try to make them resemble their living states. He proposed that the fruits contained a stigma with a funnel shaped opening in the center in which the pollen grains would get lodged.[5] The entire pollen grain would not be able to enter into the ovule, a defining trait in angiosperms. This theory was disproved 1933 by Thomas' student Tom Harris, who studied the same reproductive organs and found different results. "Most of the fruits were obtained by dissolving in hydrofluoric acid a single very small fragment of shale collected from Cape Stewart"[6] The maceration of the fruits dissolves the main cells wall and leave the cuticle and interior cell organs. This allowed Harris to look closely at the ovules located inside. Upon close inspection of the ovule, whole pollen grains were found inside the micropylar canal. This is typical of a gymnosperm reproduction, not an angiosperm. Presumably pollination was at an early stage of cupule and ovule development, before full inflation of the cupules.[1] While Thomas's original idea led many scientist to believe that Caytoniales may have been angiosperms, Harris's further research disproved this theory.
The enclosure of ovules in Caytoniales has nevertheless been considered an early stage in evolution of the angiosperm double integument, and the carpels formed from an elaboration of their stalk (Fig. 5[7]). Other theories for the origin of angiosperms derive them from Glossopteridales (Fig.5[8]), among other groups (see Evolutionary history of plants).
Gallery
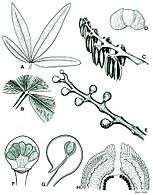 Fig. 2 A) leaf structure B) Venation C) Seed structure D) Pollen grain E) Pollen sacs F)Cupule G) Pollen sac structure H) Ovule
Fig. 2 A) leaf structure B) Venation C) Seed structure D) Pollen grain E) Pollen sacs F)Cupule G) Pollen sac structure H) Ovule Fig. 3 Fossil leaves of Sagenopteris phillipsii from the Gristhorpe Bed at Cayton Bay. Natural History Museum specimen photographed by G.J.Retallack
Fig. 3 Fossil leaves of Sagenopteris phillipsii from the Gristhorpe Bed at Cayton Bay. Natural History Museum specimen photographed by G.J.Retallack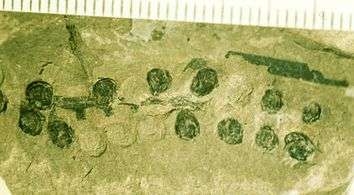 Fig. 4 Fossil Caytonia nathorstii, the reproductive structure that encloses ovules. Natural History Museum specimen photographed by G.J. Retallack.
Fig. 4 Fossil Caytonia nathorstii, the reproductive structure that encloses ovules. Natural History Museum specimen photographed by G.J. Retallack.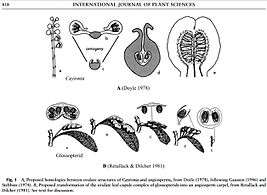
References
- Retallack, G.J.; Dilcher, D.L. (1988). "Reconstructions of selected seed ferns". Missouri Botanical Garden Annals. 75 (3): 1010–1057. doi:10.2307/2399379. JSTOR 2399379.
- Niklas, Karl J. (1997). The Evolutionary Biology of Plants. University Of Chicago Press. p. 470. ISBN 978-0-226-58083-8.
- National Academy of Sciences (2000). Ayala, Francisco J.; Fitch, Walter M.; Clegg, Michael T. (eds.). Variation and Evolution in Plants and Microorganisms: Toward a New Synthesis 50 Years after Stebbins. National Academies Press. p. 352. ISBN 978-0-309-07099-7.
- Arnold, Chester (1947). An Introduction To Paleobotany. Miller Press. p. 428. ISBN 978-1-4067-1861-4.
- Thomas, Hamshaw (1925). "The Caytoniales, A new group of angiospermous plants from the Jurassic rocks of Yorkshire". Philosophical Transactions of the Royal Society. B213 (402–410): 299–363. doi:10.1098/rstb.1925.0006.
- Harris, T.M (1933). "A new member of the Caytoniales". New Phytologist. 32 (2): 97–114. doi:10.1111/j.1469-8137.1933.tb07001.x.
- Doyle, J.A. (1978). "Origin of angiosperms". Annual Review of Ecology and Systematics. 9: 365–392). doi:10.1146/annurev.es.09.110178.002053.
- Retallack, G.; Dilcher, D.L. (1981). "Arguments for a glossopterid ancestry of angiosperms". Paleobiology. 7: 54–67. doi:10.1017/s009483730000378x.
- Friis, Else (2011). Early Flowers and Angiosperm Evolution.
- Elgorriaga, A.; Escapa, I. H.; Cúneo, R. (2019). "Southern Hemisphere Caytoniales: vegetative and reproductive remains from the Lonco Trapial Formation (Lower Jurassic), Patagonia". Journal of Systematic Palaeontology. doi:10.1080/14772019.2018.1535456.
- "Caytoniales † - Plant Evolution & Paleobotany". sites.google.com. Retrieved 2016-03-09.
- Krassilov, V.A (1977). "Contributions to the knowledge of the Caytoniales". Review of Palaeobotany and Palynology.
- "Importance of the Caytoniales". www.hullgeolsoc.co.uk. Retrieved 2016-03-09.