Calciseptine
Calciseptine (CaS) is a natural neurotoxin isolated from the black mamba Dendroaspis p. polylepis venom. This toxin consists of 60 amino acids with four disulfide bonds. Calciseptine specifically blocks L-type calcium channels, but not other voltage-dependent Ca2+ channels such as N-type and T-type channels.[1]
| Calciseptine | |
| Formula | C299H476N90O87S10 |
| Molecular weight | 7036 g/mol |
| Calciseptine | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Identifiers | |||||||
| Organism | |||||||
| Symbol | CaS | ||||||
| UniProt | P22947 | ||||||
| |||||||
History
The black mamba (Dendroaspis polylepis) is generally considered to be one of the deadliest snakes on the planet,[3] and is responsible for many fatalities throughout its sub-Saharan Africa range. Without treatment, the bite of a Black Mamba causes a 100% mortality rate.[4]
The venom of the Black Mamba consists of more than 28 peptides. One of these peptides is calciseptine. It makes up 2.8% of the venom of the Black Mamba. When first purified, the peptide was called protein E3, which was later changed to calciseptine by Weille et al.[1][4]
Importance
Calciseptine has been shown to specifically inhibit the L-type voltage-gated Ca2+ channels and was the first natural polypeptide discovered with this property. Specific polypeptide inhibitors of voltage-sensitive channels are important tools in research, and were already known for voltage-sensitive Na+ channels, both voltage-sensitive and Ca2+-activated K+ channels, and for N-type Ca2+-channels.[1]
Before calciseptine was sequenced and shown to be a specific L-type calcium channel inhibitor, no specific polypeptide inhibitors were known for this type of voltage-gated channels. Specific blockers of the L-type channel were small organic molecules like 1,4-dihydropyridines. It was suggested that polypeptide inhibitors could be found in snake venoms. Calciseptine confirmed this as it was shown to not only block the L-type channels specifically, but also to do this in exactly the same spot as the 1,4-dihydropyridines.[5] After calciseptine, other polypeptides specifically blocking the L-type channels were found as well: FS2, C10S2C2 and S4C8.[1]
Isolation/Synthesis
Calciseptine is only found in the Black Mamba and can be purified from the crude venom.[1] The snake can be milked to acquire the venom. Schweitz et al. used a three-step method to purify calciseptine to homology. The three steps were: 1) gel filtration, 2) ion exchange on TSK SP 5PW and 3) reverse-phase chromatography on RP18.[4]
Scientists also succeeded in synthesizing calciseptine in the lab. FS2, which is another component of the Black Mamba's venom and is homologous to calciseptine, can also be synthesized in the lab.[6]
Structure
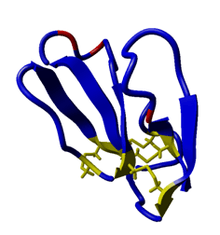
Three-fingered toxin family
Calciseptine is a member of a family of toxic peptides found in snake venoms. Proteins from this family are small and consist of 58 to 74 amino acids. They all have the same three-fingered structure which arises from the formation of four disulfide bridges between eight cysteine residues.[7]
The activities of these toxins - although they are members of the same family - are actually quite diverse and can range from the blocking of acetylcholine receptors to the changing of membrane permeability. Their common feature is that they all affect signal transduction in some way.[7]
Sequence and structure
Calciseptine itself consists of 60 amino acids and has been fully sequenced:[8]
RICYIHKASL PRATKTCVEN TCYKMFIRTQ REYISERGCG CPTAMWPYQT ECCKGDRCNK
The three-dimensional structure of calciseptine has not been determined experimentally. However, another toxin found in black mamba venom, called FS2, sequentially differs from calciseptine in only three residues: it contains a serine instead of isoleucine in position 5, a histidine instead of glutamine in position 30, and a glutamine instead of glutamic acid in position 32.[1] A three-dimensional structure of FS2 has been determined using NMR, and because of the minor sequential differences, this can serve as a model of the calciseptine structure.[9]
Toxicokinetics
Absorption and distribution
As calciseptine is injected by the snake into its prey along with the rest of its venom, it does not have to pass the protective barrier of the skin and is injected directly into the tissues and/or bloodstream of the prey. No research has been conducted on the toxicokinetics of calciseptine specifically, but general research about snake toxin dynamics is available. Although the toxic peptides are generally small (about 60 amino acids), their size is sufficient to prevent them from crossing epithelial layers like the blood-brain barrier.[10]
Bioavailability measurements have been conducted for several snake venoms. For example, cobra venom has been found to have a bioavailability of 41.7% when injected intramuscular, and for other venoms this may even be less than 10%. These values are quite low compared to those of most therapeutic drugs, which usually have a bioavailability of nearly 100% after intramuscular injection.[11] In general, toxic peptides of 10-40 amino acids have been found to have a relatively poor bioavailability due to their size and hydrophilicity.[10] Thus, calciseptine, containing 60 amino acids, is expected to have a low bioavailability as well.
Metabolism
Because calciseptine is a peptide, theoretically it can be broken down by proteases in the tissues where it is injected. It has been found that digestion of snake toxic peptides by proteases does occur in the prey tissues, but due to the relative stability of the toxins, the speed with which the toxins act and the amount of venom injected, this is not enough to protect against the consequences of a snake bite. The same goes for the immune system: the larger venom peptides are unlikely to be missed by the immune system, but immunological action is not fast enough to counter the effects of the venom.[10][12]
As mentioned above, calciseptine has a three-fingered structure commonly found in toxins. Although many toxins have this structure, their functions are diverse. The reason this structure is so much conserved is probably its stability: the cysteine bridges create a stable core, which possibly slows the breakdown of the protein by proteases.[10]
Mechanism of action
Calciseptine has been shown to block L-type calcium channels, thus inhibiting smooth muscle contraction and cardiac function.[1]
Voltage-activated calcium channels
Voltage-dependent calcium channels are important for generating electrical signals in excitable cells like neurons and cardiac or smooth muscle cells. N-type Ca2+ channels are found in neuronal cells, and play an important role in the coupling of nerve excitation and neurotransmitter secretion. L-type calcium channels are present in cardiac and smooth muscle cells, coupling excitation to muscle contraction. Other types of voltage-activated Ca2+-channels include T-type and P-type channels.[1]
Blocking of L-type calcium channels
Because the presence of calcium in the cytosol is required for muscle contraction, blockers of calcium channels prevent the muscles from building tension. Blockers of the L-type calcium channels, which occur in cardiac and smooth muscles, thus act as smooth muscle relaxants and inhibitors of cardiac contractions.[1] Common blockers of L-type calcium channels are 1,4-dihydropyridines, which are used in treatment of cardiovascular diseases. Because of their activity as a relaxant, they can relax smooth muscles surrounding blood vessels, thus widening them and lowering the blood pressure.[1][7][13]
Calciseptine as an L-type calcium channel blocker
Calciseptine resembles the abovementioned 1,4-dihydropyridines in its biological action, as it has the same ability to bind and block the L-type calcium channels in smooth and cardiac muscle.[1]
The amino acids responsible for binding and blocking the L-type calcium channels are probably located in the third ‘finger’ of the calciseptine structure, somewhere between amino acids 40 and 50. Although peptides from the three-fingered family are alike in structure, only some of them are able to bind and block calcium channels. Multiple sequence alignment studies yielded 12 amino acid residues that were unique to the toxins with channel-blocking activities. These residues are located at the tips of loops II and III in the three-fingered structure. A model has been proposed in which the amino acids 45 to 48, MWPY, of the FS2 toxin are considered to bind the calcium channels. This model is based on a prediction of the interactions of these amino acids with the calcium channel. These interactions resemble the hydrophobic and hydrogen bonding properties of nifedipine, a known 1,4-dihydropyridine blocker.[7] Because of the similarities between calciseptine and FS2, this model could account for the interactions of calciseptine with the L-type calcium channels as well.
Another model is based on a larger segment of the same loop, containing amino acids 42 to 47, PTAMWP. Because sites of protein-protein interaction have often been found to be flanked by prolines,[14] the presence of the two prolines in this ‘finger’ of the structure indicates a possible interaction site. A short polypeptide of eight amino acid residues, containing this sequence, was indeed found to block the L-type calcium channels, though with a lesser activity.[15]
These interacting amino acids are also found in two other L-type calcium channel blocking proteins, C10S2C2 and S4C8. Although both of these toxins are members of the three-fingered family, the three-fingered structure is probably not a requirement for channel blocking: other snake toxins, including dendrotoxin, have similar effects on various channels, but do not show the three-fingered structure.[7][15]
Indications/Symptoms
Typical symptoms after being bitten by a Black Mamba include the rapid onset of dizziness, drowsiness and coughing and having difficulties breathing. Other likely symptoms include convulsions, neuromuscular symptoms, shock, loss of consciousness, hypotension, ataxia, excessive salivation, limb paralysis, nausea and vomiting, fever, and severe abdominal pain. Permanent limb paralysis is very likely if the bite remains untreated. In the most severe case, if untreated, the bite of the Black Mamba can lead to death by suffocation, resulting from the paralysis of respiratory muscles.[16]
All these symptoms are due to a combination of all the toxic peptides the crude venom of the Black Mamba contains. The symptoms are related to calciseptine because it also works as a smooth muscle relaxant,[1] thus explaining the early onset of having difficulties breathing, limb paralysis and even death by suffocation. Calciseptine is known to have a hypotensive effect.[17] It cannot clearly be established which peptide is most responsible for which symptom, since combinations of different toxins can have various effects.
Antidote
In case of a bite from the black mamba, the victim should be treated according to a standard protocol. The most important part of this treatment is the intravenous injection of a polyvalent antivenom. South African Vaccine Producers produces this antivenom. Polyvalent means that it can be used for different snakebites: vipers, mambas and cobras.[18] Large quantities of the antivenom must be injected to counter the effects of the venom.
The polyvalent antivenin is produced by injecting horses with adapted venom. The venom is first detoxified to prevent too much damage and death. This is mostly done by complexing the venom with an aldehyde like formalin. The venom is also administered with an adjuvant, like aluminium hydroxide or sodium alginate, to stimulate the immunological response. When the venom is injected, the body will produce antibodies. These will bind components – the variability of peptides – of the venom, which prevent further activity of the molecule and are ultimately removed by the immune system of the body. These antibodies are collected and purified from the blood and then packaged in mostly a liquid form. Horses are used because of the large blood volume. The final antivenom product expires after 5 years and needs to preferentially be kept cool, 4-8 °C. It can, however, survive different environmental situations for some weeks to months, without losing its potency.[19]
Because several venoms of different snakes are injected over time, the horse will develop different antibodies against all the venoms. This gives the antivenin its polyvalent property. This also enhances the potency of the antivenin. Some venoms cannot induce the immune system enough on its own or the combination of two or more venoms result in a better responding antivenin.[19]
Toxicity
Lethality
Calciseptine is toxic in low doses. No specific data are available on the toxicity of calciseptine in humans. However, the LD50 values for mice have been determined and can be found in the table below:[20]
| Mode of administration | LD50 value |
|---|---|
| Intravenous injection | 0.25 mg/kg |
| Subcutaneous injection | 0.38 mg/kg |
| Intraperitoneal injection | 0.94 mg/kg |
Venom toxicity
The Black Mamba can inject between 100–120 mg venom in one bite. Comparing the LD50 results with the amount of venom that can be injected shows that one bite is fatal. For humans a dose of 10–15 mg is enough to be fatal.[21]
Other data
Calciseptine has an effect on the K+-induced contractions and L-type Ca2+ channel activity, the IC50 values were determined and gave values of 230 and 430 nM respectively. Calciseptine also has an effect on the cardiac function. The IC50 value found for this tissue was 15 nM. These values were determined in rats and mice.[1]
Synergistic toxicity
It has been found that some three-fingered toxins from mamba venom interact synergistically with each other. The molecular mechanisms of these interactions remain unknown.[22] It is not known either whether calciseptine has a synergistic effect with other venom compounds.
Effects on Animals
Calciseptine has been examined in vivo and in vitro in all kinds of animals, but mostly in rats. Calciseptine relaxes precontracted rat (thoracic) aorta and decreases blood pressure drastically.[17]
The decrease in blood pressure shows a combination of short- and long-lasting effects. The early, acute onset took five minutes, and the effect could last for 120 minutes or longer. In addition, calciseptine had only a small effect on the heart rhythm, changing it only slightly. Furthermore, it can also relax the trachea rings in the lungs. These effects can be explained by the relaxing activity of calciseptine on various smooth muscle cells.[1] The inhibitory effect of calciseptine results in a decreased or total disappearance of electric activity in these cells. The total inhibitory effect depends on the tissue: the cardiovascular system is the most vulnerable, while neuronal cells are less vulnerable and skeletal muscle cells are completely resistant. This difference in tissue sensitivity is probably caused by slight differences in the L-type calcium channels in these tissues.[1] These effects can occur at low subjected amounts of 0.1 till 1 μM calciseptine. In mouse myotube the Ca2+ currents show higher amplitude after incubation in calciseptine (1 μM). This effect of calciseptine on the Ca2+ current develops relatively fast. Calciseptine changes the reversal potential of the Ca2+ current in mouse myotubes.[23] In adult frog skeletal muscle fibers calciseptine also causes an increased Ca2+ current. This increase is comparable to the increase which was found in mouse myotubes. In contrast to the mouse myotubes calciseptine did not change the reversal potential of the Ca2+ current. In rat muscle fibers after applying calciseptine there were very slight changes found in twitch tension, which shows us that the peptide had very little effect on muscle contraction. Calciseptine also had little effect on frog muscle fibers.[23]
There is a small effect of calciseptine during a series of repetitive stimuli which generate a tetanus; calciseptine increased tetanic tension. The average tetanic tension after applying calciseptine was slightly higher than the control value. Though calciseptine gives an increase in charge movement and increase in influx of Ca2+ through L-type channels this has no major effects on contraction of the muscle, even in tetanus.[23]
Research on rats and guinea pigs revealed that synthetic calciseptine and FS2 as well, have the same effect as their natural counterparts.[17]
References
- De Weille J. R., Schweitz H., Maes P., Tartar A., Lazdunski M (1991). "Calciseptine, a peptide isolated from black mamba venom, is a specific blocker of the L-type calcium channel". Proc. Natl. Acad. Sci. USA. 88 (6): 2437–2440. Bibcode:1991PNAS...88.2437D. doi:10.1073/pnas.88.6.2437. PMC 51247. PMID 1848702.CS1 maint: multiple names: authors list (link)
- Haji, R. "Venomous snakes and snake bite" (PDF). Zoocheck Canada Inc. Archived from the original (PDF) on 25 April 2012. Retrieved 2 December 2011.
- Schweitz H, Bidard JN, Lazdunski M (1990). "Purification and Pharmacological Characterization of Peptide Toxins from the Black Mamba (Dendroaspis Polylepis) Venom". Toxicon. 28 (7): 847–856. doi:10.1016/s0041-0101(09)80007-x. PMID 1977212.
- Yasuda O, Morimoto S, et al. (1993). "Calciseptine binding to 1,4-dihydropyridine recognition site of the L-type calcium channel of rat synaptosomal membranes". Biochemical and Biophysical Research Communications. 194 (2): 587–594. doi:10.1006/bbrc.1993.1862. PMID 8393668.
- Kuroda, H.; et al. (1992). "Solution Synthesis of calciseptine, an L-type specific calcium channel blocker". Pept. Res. 5: 265–268.
- Schleifer, K. (1997). "Comparative molecular modelling study of the calcium channel blockers nifedipine and black mamba toxin FS2". Journal of Computer-Aided Molecular Design. 11 (5): 491–501. Bibcode:1997JCAMD..11..491S. doi:10.1023/a:1007974124426. PMID 9385552.
- "Uniprot KB". Retrieved 3 April 2013.
- Albrand, J.P.; et al. (1995). "NMR and restrained molecular dynamics study of the three-dimensional solution structure of toxin FS2, a specific blocker of the L-type calcium channel, isolated from black mamba venom". Biochemistry. 34 (17): 5923–5937. doi:10.1021/bi00017a022. PMID 7727450.
- Lewis, R.J.; Garcia, M.L. (2003). "Therapeutic potential of venom peptides". Nature Reviews Drug Discovery. 2 (10): 790–802. doi:10.1038/nrd1197. PMID 14526382.
- Yap, M.K.K.; et al. (2013). "Toxicokinetics of Naja sputatrix (Javan spitting cobra) venom following intramuscular and intravenous administrations of the venom into rabbits". Toxicon. 68: 18–23. doi:10.1016/j.toxicon.2013.02.017. PMID 23537711.
- Bottrall, J.L.; et al. (2010). "Proteolytic activity of elapid and viperid snake venoms and its implication to digestion". J Venom Res. 1: 18–28. PMC 3086185. PMID 21544178.
- Varon, J.; Marik, P.E. (2003). "Clinical review: The management of hypertensive crises". Crit. Care. 7 (5): 374–384. doi:10.1186/cc2351. PMC 270718. PMID 12974970.
- Kini, R.M. (1998). "Proline brackets and identification of potential functional sites in proteins: toxins to therapeutics". Toxicon. 36 (11): 1659–1670. doi:10.1016/s0041-0101(98)00159-7. PMID 9792183.
- Kini, R.M.; et al. (1998). "Flanking proline residues identify the L-type ca2+ channel binding site of calciseptine and FS2". Biochemistry. 37 (25): 9058–9063. doi:10.1021/bi9802723. PMID 9636051.
- "Immediate first aid". Archived from the original on 2012-11-02. Retrieved 2013-03-27.
- Watanabe, T.; et al. (1995). "Smooth Muscle Relaxing and Hypotensice Activities of Synthetic Calciseptine and the Homologous Snake Venom Peptide FS2". Jpn. J. Pharmacol. 68 (3): 305–313. doi:10.1254/jjp.68.305. PMID 7474554.
- "African Reptiles & Venom". Archived from the original on 8 May 2013. Retrieved 3 April 2013.
- Chippaux, J.P.; Goyffon, M. (1998). "Venoms, antivenoms and immunotherapy". Toxicon. 36 (6): 823–846. doi:10.1016/s0041-0101(97)00160-8. PMID 9663690.
- "Toxin and Toxin Target Database". Retrieved 3 April 2013.
- Branch, William R. (1998). Field Guide to Snakes and Other Reptiles of Southern Africa. Cape town: Struik Publishers.
- Kini, R.M.; Doley, R. (2010). "Structure, function and evolution of three-finger toxins: Mini proteins with multiple targets". Toxicon. 56 (6): 855–867. doi:10.1016/j.toxicon.2010.07.010. PMID 20670641.
- Garcia, M.C.; et al. (2001). "Calciseptine, a Ca2+ Channel Blocker, has Agonist Actions on L-Type Ca2+ currents on Frog and Mammalian Skeletal Muscle". J. Membrane Biol. 184 (2): 121–129. doi:10.1007/s00232-001-0080-7. PMID 11719849.