Racialism
Racialism is the idea that humanity can be easily divided into well-defined biological categories ("races") that are both broad (each category should include many humans, such as entire continents' populations) and clearly-defined (the categorization method should rarely misidentify someone's "race"). Racialism implies that these races are substantially physiologically different from each other and that these racial differences strongly determine the abilities and behavior of individuals and peoples. Essentially, racialism argues that human populations are substantially different from each other to a degree which necessitates biological classification below the species level. In short, racialism holds that ethnicity has a biological component in addition to the sociocultural one.
| The colorful pseudoscience Racialism |
| Hating thy neighbour |
| Divide and conquer |
| Dog-whistlers |
|
v - t - e |
“”Race is not an accurate or productive way to describe human biological variation. |
| —Edgar et al. 2009[1] |
This Pseudoscience should not be confused with Racism, which is the belief that one or more of these various groups is "superior" or "inferior" to one or more others, usually in some vague, undefined way. While Racism and Racialism are distinct concepts, their respective adherents do overlap a lot more than either would have you believe.
Very few racialists call themselves "racialist". Their preferred labels include scientific racism (pre–2000),[note 1] race realism or racial realism (post–2000),[note 2] and human biodiversity or HBD (post–2010).[note 3]
Racialism was first developed in the 1700s. It remained virtually unchallenged until the 1930s-1960s, when genetics showed it to be erroneous. Simply put, modern racialism is pseudoscientific bullshit.[1][2][3][4]
Human biological variation
- To deny the existence of subspecies in Homo sapiens is not to deny biological variation between populations; there is human population structure.
- Misrepresenting anti-racialism as the position that "all human populations are identical" or there are "no differences between populations" is a common tactic among racialists.
Populations (demes)
Human individuals (of opposite sexes) do not randomly reproduce because of geographical constraints: "panmixia
Demes should not be confused with races.[9] Typically, demes are very small and localised breeding near-isolates, including tribes, rural village populations and ethno-religious groups such as the Lancaster Amish,[10] that have a very high rate of endogamy, although minimal gene flow still exists, somewhat blurring the boundaries of each deme:
Deme (pronounced “deem,” from the Greek for “people” and originally referring to a political division within ancient Greece) has been used in biology since the 1930s as a term for a local interbreeding population within a species. As such, the recognition of demes can be confused with, and can appear to provide justification for, the existence of biological races or subspecies.
The rationale for naming population units below the species level comes from the simple fact that members of a species are seldom, if ever, evenly distributed throughout the species’ geographical range. Uneven distribution can result in clusters of individuals partially isolated from other such clusters—that is, with more interbreeding within the clusters than between them—simply because of proximity. It is to such clusters that the term deme is usually applied. Thus, the green frogs in an isolated pond, a town of prairie dogs, or a field of wild sunflowers might be examples of demes. If demes inhabit different local environments, natural selection can operate in different directions in these populations with the result that there may be genetic and even physical variation in the characteristics of individual demes. Other processes of evolution, such as mutation and other forms of genetic change, can also enhance these differences, depending upon the extent of demic isolation.
The problem with the concept of the deme is that there is no definitive set of criteria for recognizing demes within species. Normally, some spatial separation or other obvious impediment to genetic exchange is a clue, with genetic or physical distinctions as an expected result. Demes, however, are populations within species, and by definition, exchange genes with other demes of the same species either directly, in the case of adjacent demes, or by a series of steps, in the case of widely separated demes.
Demes are also transitory. According to Stephen Jay Gould in The Structure of Evolutionary Theory, demes have “porous borders,” they do not function as a “discrete [evolutionary] entities,” and are “defined only by the transient and clumpy nature of … habitats” (p. 647). Richard Dawkins, in The Selfish Gene, likened demes to “clouds in the sky or dust storms in the desert … temporary aggregations” (p. 36). Moreover, even if demes are spatially distinct clusters, their phenotypic features might still grade into one another in response to environmental gradients, thus making the boundaries between the demes even less distinct. This is known as a clinal distribution.
Given the accepted general definition, it can certainly be said that demes exist within the human species, where they are identified as semi-isolated endogamous (literally, “marriage within”) populations. The isolation may be spatial, as in any of a number of societies inhabiting separate and secluded valleys in the New Guinea Highlands, or cultural, as in the religiously based isolation of groups such as the Hutterites, Mennonites, and Amish.
All human populations consist of members of a single species. Thus, by definition, they exchange genes with other populations, directly or indirectly. More specifically, even the most isolated human population has experienced gene flow with adjacent populations at some point in time. Outsiders have always married into the Hutterites, for example, and Hutterites have married out. Over time, human demes are as ephemeral as those of any other species, and probably more so, given the human proclivities for mobility and genetic exchange.
The question becomes, then, what is the relationship between the concept of the deme and that of the biological race or subspecies? Both are proposed populations below the species level. Traditionally, a biological race was considered to be a group of related demes inhabiting the same general region and sharing genetic and/or physical characteristics. The limitations noted for the deme concept, however, provide a lesson for considering such larger groups as races or subspecies because those limitations become more pronounced with larger populations. Larger populations within a species contain more genetic and physical variation, and thus stand even less chance of having specific biological characteristics. Because there is more gene flow between large populations, the boundaries between such populations are further blurred and have less real biological meaning and utility.[11]
Population genetics
The persistence of geographical and/or cultural barriers between demes over many generations has led to genetic (and phenotypic) differences. The biologist Jonathan Marks describes these populations as the "small biopackages" of human variation, but cautions that overall differences between populations are small, hence his position is "populations are biologically real, not races".[12] In population genetics, these differences are analysed by mean variation in a large number of allele (variant of a gene) frequencies, or a smaller number of AIMs
In their publication The History and Geography of Human Genes, Cavalli-Sforza, Menozzi & Piazza (1994) include genetic samples from 491 breeding populations from across the globe; the authors note that "changes in means of transportation and labour opportunities have of course altered profoundly marriage customs" and found that "an increase in geographic distance between birth of mates" somewhat complicated their study. More recently, Cavalli-Sforza (2005) has worked on the Human Genome Diversity Project that includes genetic samples from 52 ethnic groups from different continents.[13] The status these ethnic groups as demes is questionable because of their larger size in terms of numbers of individuals and more vague boundaries. Cavalli-Sforza recognises this issue; for example Han Chinese, who number more than a billion individuals and who live across the entirety of China, he does not consider a single breeding population. In the case HGDP samples from small-size ethnic groups restricted to local areas, e.g. Suruí
The biologist John Relethford throughout his work stresses that population genetics should focus on micro-size populations because these reflect actual mating behaviours; a larger number of individuals across more space is likely to contain different breeding populations. As explained by Jonathan Marks in a 2010 article: "larger units than the deme lack cohesion... to adopt a unit of analysis of human biology larger than that of the local population or deme, then, is what requires some justification today".[14] Like Relethford and Cavalli-Sforza et al., Marks goes on to deny the validity of aggregating demes into too broad divisions (large countries and continents); firstly since their boundaries are too arbitrary, and secondly because they have too many individuals (with different tendencies of assortative mating or endogamy) and so they are not useful. Relethford (2017) concludes by saying:
Application of much of population genetics works best when considering variation between local populations and not between aggregates. The fine detail of our species' evolutionary history and its impact on patterns of genetic variation are lost when trying to categorize and classify into races. The proper study of human biological variation needs to be rooted in evolutionary theory and population genetics.[15]
Clines
“”There are no races, there are only clines. |
| —Frank B. Livingstone, 1962[16] |
Interpopulation variation is mostly continuous, and the arbitrary categorization of humans into races is therefore neither useful nor an accurate method in analyzing biological variation. Instead, modern biologists and physical anthropologists tend to study clines.
Clines replaced the race concept from the 1960s.[17] In their work Race or Clines?, Lieberman & Rice (1996) write that the rise of the cline concept was one of several facts that "laid low the concept of race".[18]
The word "cline" derives from the Greek word for slope and the smooth gradient or continuity of a given trait across geographic space is known as a clinal distribution. As Lieberman 2003 explains:[2]
Strong genetic influence weakening the race concept is also seen growing out of the concept of clines and supporting data. Clines referred to gradations of genotypes or phenotypes over a geographic area, and were distributed within and across racial and national boundaries.
Scientific viewpoint
Modern scientific consensus is strongly against the race concept, though this was not always the case. In contrast, biological evidence for racialism has always been relatively weak, and modern evidence conclusively shows that racialism is fundamentally flawed.
Scientific consensus: Races aren't useful
“”[B]oundaries in global variation are not abrupt and do not fit a strict view of the race concept; the number of races and the cutoffs used to define them are arbitrary. The race concept is at best a crude first-order approximation to the geographically structured phenotypic variation in the human species. |
| —biological anthropologist John Relethford in 2009[19] |
Major institutions with expertise have come down against the biological usefulness of race:
“”DNA studies do not indicate that separate classifiable subspecies (races) exist within modern humans. While different genes for physical traits such as skin and hair color can be identified between individuals, no consistent patterns of genes across the human genome exist to distinguish one race from another. There also is no genetic basis for divisions of human ethnicity. |
| —Human Genome Project[20] |
“”For centuries, scholars have sought to comprehend patterns in nature by classifying living things. The only living species in the human family, Homo sapiens, has become a highly diversified global array of populations. The geographic pattern of genetic variation within this array is complex, and presents no major discontinuity. Humanity cannot be classified into discrete geographic categories with absolute boundaries. Furthermore, the complexities of human history make it difficult to determine the position of certain groups in classifications. Multiplying subcategories cannot correct the inadequacies of these classifications. |
| —American Association of Physical Anthropologists[21] |
“”With the vast expansion of scientific knowledge in this century, however, it has become clear that human populations are not unambiguous, clearly demarcated, biologically distinct groups. Evidence from the analysis of genetics (e.g., DNA) indicates that most physical variation, about 94%, lies within so-called racial groups. Conventional geographic "racial" groupings differ from one another only in about 6% of their genes. This means that there is greater variation within "racial" groups than between them. In neighboring populations there is much overlapping of genes and their phenotypic (physical) expressions. Throughout history whenever different groups have come into contact, they have interbred. The continued sharing of genetic materials has maintained all of humankind as a single species. |
| —American Anthropological Association[22] |
Scientific consensus follows the above statements. Lieberman and Reynolds first attempted to determine scientific consensus on the concept of race in 1978.[23] The number of racialist researchers has continuously declined since then.[24] This can easily be seen in the decline of papers using the race concept. For example, the percent of variation articles in the American Journal of Physical Anthropology that use the race concept dropped from ~70% in the 1910's to ~15% in the 1990's.[2]

A 2012 survey of 3,286 American Anthropological Association members found very strong agreement that the conventional concept of race is not scientifically useful.[note 4] The first 10 of questions are reproduced below:[3][note 5]
| Anthropologists' views on race, ancestry, and genetics | ||||
|---|---|---|---|---|
| Statement | Most common response | Correlation of response and familiarity with genetic ancestry inference[note 6] | N | |
| The human population may be subdivided into biological races. | Strongly disagree or disagree (86%) | 0.11 (<.00010) | 1,909 | |
| Racial categories are determined by biology. | Strongly disagree or disagree (88%) | 0.11 (<.00010)* | 1,903 | |
| There are discrete biological boundaries among races. | Strongly disagree or disagree (93%) | 0.14 (<.00010)* | 1,868 | |
| Biological variability exists but this variability does not conform to the discrete packages labeled races. | Strongly agree or agree (89%) | −0.035 (.12) | 1,894 | |
| Boundaries between what have been called races are completely arbitrary, depending primarily upon the wishes of the classifier. | Strongly agree or agree (69%) | 0.0014 (.95) | 1,898 | |
| Continental population categories — Africans, Asians, Europeans — are the same as standard anthropological racial classifications. | Strongly disagree or disagree (73%) | 0.070 (.0025)* | 1,872 | |
| Continental population categories — Africans, Asians, Europeans — are useful for examining genetic relationships (i.e., relatedness) among people. | Strongly disagree or disagree (38%) | 0.21 (<.00010)* | 1,879 | |
| Race — as defined by the U.S. Office of Management and Budget (i.e., census categories) — is a useful proxy for ancestry. | Strongly disagree or disagree (73%) | 0.043 (.057) | 1,890 | |
| Genetic ancestry — inferred from genetic markers — rather than race, is a better proxy for genetic relationships among sub-Saharan Africans, Asians, Europeans, Pacific Islanders, and Native Americans. | Strongly agree or agree (75%) | 0.18 (<.00010)* | 1,887 | |
| The distributions of physical traits overlap among races. | Strongly agree or agree (89%) | 0.016 (.48) | 1,883 | |
It should be clear that anthropologists disagree with the race concept. They do so because of anthropological and genetic evidence, presented below.
Background: Comparison with animals
There are two major scientific definitions of race in animals, as Templeton 2013 explains.[25] The first is "geographically isolated & genetically different subpopulations":
One definition regards races as geographically circumscribed populations within a species that have sharp boundaries that separate them from the remainder of the species (Smith, Chiszar, & Montanucci, 1997). In traditional taxonomic studies, the boundaries were defined by morphological differences, but now these boundaries are typically defined in terms of genetic differences that can be scored in an objective fashion in all species. Most demes or local populations within a species show some degree of genetic differentiation from other local populations, by having either some unique alleles or at least different frequencies of alleles. If every genetically distinguishable population were elevated to the status of race, then most species would have hundreds to tens of thousands of races, thereby making race nothing more than a synonym for a deme or local population. A race or subspecies requires a degree of genetic differentiation that is well above the level of genetic differences that exist among local populations. One commonly used threshold is that two populations with sharp boundaries are considered to be different races if 25% or more of the genetic variability that they collectively share is found as between population differences (Smith, et al., 1997). A common measure used to quantify the degree of differentiation is a statistic known as pairwise fst. The pairwise fst statistic in turn depends upon two measures of heterozygosity. The frequency with which two genes are different alleles given that they have been randomly drawn from the two populations pooled together is designated by Ht, the expected heterozygosity of the total population. Similarly, Hs is the average frequency with which two randomly drawn genes from the same subpopulation are different alleles. Then, fst=(Ht-Hs)/Ht. In many modern genetic studies, the degree of DNA sequence differences between the randomly drawn genes is quantified, often with the use of a model of mutation, instead of just determining if the two DNA sequences are the same or different. When this done, the analysis is called an Analysis of MOlecular VAriation (AMOVA), and various measures of population differentiation analogous to fst exist for different mutation models. Regardless of the specific measure, the degree of genetic differentiation can be quantified in an objective manner in any species. Hence, human races can indeed be studied with exactly the same criteria applied to non-human species. The main disadvantage of this definition is the arbitrariness of the threshold value of 25%, although it was chosen based on the observed amount of subdivision found within many species.
Note: Templeton misrepresents Smith 1997 on the 75% rule,[26] which refers to phenotypic rather than genetic differences.[27][28] There's no hard-set rule for the exact FST cutoff, though 25% is commonly used.[29][30][31][32]
The second is "evolutionarily independent subpopulations":
A second definition defines races as distinct evolutionary lineages within a species. An evolutionary lineage is a population of organisms characterized by a continuous line of descent such that the individuals in the population at any given time are connected by ancestor/descendent relationships. Because evolutionary lineages can often be nested together into a larger, more ancestral evolutionary lineage, the evolutionary lineages that are relevant for defining subspecies in conservation biology are the smallest population units that function as an evolutionary lineage within a species. The phylogenetic species concept elevates all evolutionary lineages to the status of species (Cracraft, 1989), but most species concepts allow for multiple lineages to exist within a species. For example, the cohesion species concept defines a species as an evolutionary lineage that maintains its cohesiveness over time because it is a reproductive community capable of exchanging gametes and/or an ecological community sharing a derived adaptation or adaptations needed for successful reproduction (Templeton, 1989, 2001). Two or more evolutionary lineages nested within an older lineage that are capable of exchanging gametes and/or share the same adaptations necessary for successful reproduction are considered lineages nested within a single cohesion species. The biological species concept only uses the criterion of gamete exchangeability and is a proper logical subset of the cohesion concept (Templeton, 1998b; Templeton, 2001). Hence, the biological species concept also allows multiple evolutionary lineages to exist within a species. The possibility of multiple evolutionary lineages within a species is commonly recognized in the area of conservation biology, and indeed the evolutionary lineage definition of race or subspecies has become the dominant definition in much of conservation and evolutionary biology, in large part because it is a natural historical population unit that emerges from modern phylogenetic theory and practice (Amato & Gatesy, 1994; Crandall, Binida-Emonds, Mace, & Wayne, 2000).
Both of these concepts are rejected within the human species.[25]
Definition One: Human gene clusters don't divide neatly into geographical groupings
“”If human races are geographically delimited populations characterized by regionally distinctive phenotypes that do not occur elsewhere in significant numbers, then races no longer exist and have probably not existed for centuries, if ever. |
| —Cartmill 1998[33] |
Human genetics doesn't work like racialists think it does. Race realists spend a great deal of time and effort pointing out genetic differences between geographically separated populations in gene clustering research and insisting this is evidence for "races".
In scientific work, gene clustering research in a set of populations is typically determined via subjective descriptors of ethnicity, language and geography. In this way, people can be reliably identified as members of these groups.[34] However, this way of categorizing people depends fundamentally on the quantity and method used to create the aforementioned framework of ancestral populations; how people are grouped into populations is completely arbitrary.[34]
In contrast, racialism relies upon the idea that large clusters of people who are homogeneous within the cluster and heterogeneous between clusters in terms of genetic similarity exist. However, this idea has no scientific basis and in fact there is evidence against it, as Witherspoon et al. concluded in their 2007 paper "Genetic Similarities Within and Between Human Populations".[35] As Maglo et al. 2016 write:[34]
An evolutionary-based grouping of world populations attempts to summarize the complex human population history while an instrumental grouping lumps pragmatically world populations into five continental groups reducing evolutionary relations. It is this instrumentally engineered clustered picture of human evolutionary history that is misleadingly construed as corresponding to socially defined races in countries such as the US. Although these socially defined races and continental genetic clusters do not actually match, the alleged correspondence has generated its own sets of debates.
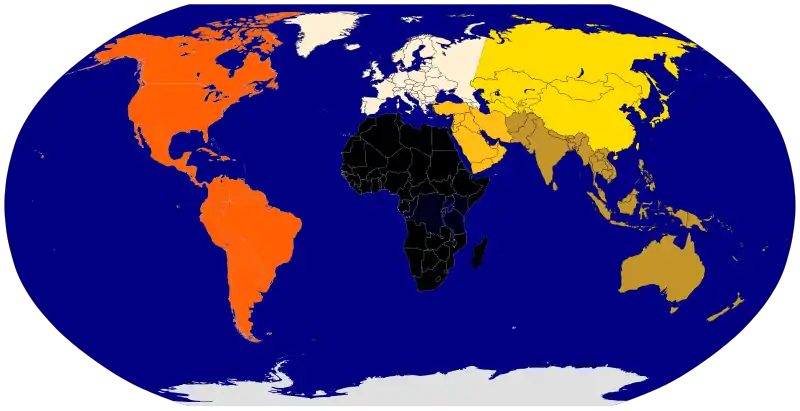
"Clusters" of very high genetic similarity can be -- purposefully or accidentally -- "created" via inadequate sampling of intermediate populations. In this case, three "races" are obtained (blue, purple, red) even though the overall system shows no such breaks (Maglo et al. 2016):[34]

The above distinctions should be made clear when comparing a racialist map of the world (left), the scientific clinal map of the world (center), and human Y-chromosome haplogroups

Genetic evidence makes it readily apparent that there are no sudden "breaks" in human variation. Instead, there are merely gradations ("clines"). Otherwise, it should be simple enough to point out substantial breaks in the following graphs of genetic variation of the human species:[37]


In short: There are no sharply defined "jumps" of genetic difference between humans, and so no reason to have sharply defined races of humans.
Rosenberg et al. 2005
A study often cited by racialists as evidence there are continental races is Rosenberg et al. 2005.[38] Rosenberg and colleagues claimed to have discovered a very small percentage of genetic variation (1.53%) is found between continents, i.e. "discontinuous jumps in genetic distance—across oceans, the Himalayas, and the Sahara". However their study only used 52 population samples. Notably no samples were included from the Sahara nor Himalayas, therefore it is not a surprise their study erroneously found genetic discontinuity between continents by poor sampling:
The importance of this point is testified to by the fact that Rosenberg and colleagues confirmed yet again that ‘intermediate’ populations for which there were samples—such as in Pakistan and western China—did not reveal [allele] discontinuities between the populations they ostensibly separate, a result that likely holds globally." (Joshua Glasgow, A Theory of Race
File:Wikipedia's W.svg , 2009, p. 106)
With a lot more population samples, Tishkoff et al. (2009) found no genetic discontinuity between the Saharan desert and Sahel, instead a genetic continua runs "across northern Africa into Mali (the Dogon)"[39] (see also Serre & Pääbo, 2004).[40]
Definition One: Human phenotypes don't divide neatly into geographical groupings
“”And if races are not geographically delimited, then racial classificatory categories are merely labels for polymorphisms that vary in frequency from one part of the world to another, like redheadedness or Type A blood. If "Negroid" and "Caucasoid" people occur on every continent, it makes no more sense to describe these groupings as geographical subspecies than it would to describe redheads or people with Type A blood as human subspecies. |
| —Cartmill 1998[33] |
Expressed genes turn into phenotypes, the actual manifestation of differences. Phenotype is, at times, fairly close to genotype and, thus, might fit into the genetics of race. However, it too is a flawed metric.
As a case study: The highest level of long-distance runners comes not just from Kenya, but a particular part of Kenya and their success is defined, in part, by common genetics.[41] But that isn't race. In fact, the opposite end of the running spectrum, sprinters, features a large proportion with ancestral origins in West Africa.[42][43] Both sets of genes favor running, but entirely different aspects of it. By these genes alone, the two groups might appear to be different races, even while our knee-jerk appearance-based definitions put them in the same race.
Applying the same technique as Witherspoon,[35] Strauss and Hubbe analyzed data of head shape and found that, across three regions that correspond to three common racial definitions, a typical person in any region was about 30% likely to be more similar to someone from a different region than to someone within their own region.[44] In other words, head shape, one defining factor of race, does not correspond consistently to geographical heritage in the way it would be expected were genetic differences determinant of race. In even shorter words, the genotype may be similar among a race, but phenotype is less similar. This may explain how one white supremacist was so surprised to discover his African heritage.[45]
Definition Two: Human races are not evolutionarily independent
Racialist views oversimplify the complicated history of humanity. For example, the racialist tree diagram



Genetic evidence: Race does not predict human variation well
If race were more than just a social construct, the genetics of race should show differences strongly associated with the definitions of race. The easiest distinctions come from things like skin color, hair, and eye shape which, yes, are genetically based, but the surface commonality is not enough to justify racial stereotypes. For those stereotypes to be true, the genetics of race have to extend deeper.
However, modern genetic data has demonstrated the vast majority of variation (>85%) is found within populations, rather than between them (<15%).[46][47][48][49][50] Of the latter, <10% is found between large continental groups of ethnic groups/demes (such as "white" or "black" or "Asian").
It is possible to compare the variation of various phenotypic markers (eg, skull structure) genetic markers (ie, DNA patterns) within populations and between populations. The vast majority of these studied differences have yielded low rates of deviation between races and between ethnic groups. This suggests that the vast majority of deviation actually occurs within populations, rather than between populations. In turn, this casts serious doubt on the term "race" having any meaning whatsoever -- if the vast majority of variation is within populations, then it makes little sense to categorize each population. As the below table of scientific studies of differences between human population groups (Taken in part from Ruvolo & Seielstad 2001[4]) should show, it is the case that the vast majority of variation occurs within populations.
| Neutral genetic/phenotypic markers | ||||
|---|---|---|---|---|
| Data | Reference | Percent of deviation within populations | Percent of deviation between ethnic groups | Percent of deviation between "races" |
| 17 classical polymorphisms[note 8] | Lewontin (1972)[51] | 85.4% | 8.3% | 6.3% |
| 18 classical polymorphisms[note 9] | Latter (1980)[52] | 85.5% | 5.5% | 9.0% |
| 25 classical polymorphisms[note 10] | Ryman et al. (1983)[53] | 86.0% | 2.8% | 11.2% |
| 57 landmarks and dimensions of the skull | Relethford (1994,[54] 2002[55]) | 78.8% | 6.7% | 14.6% |
| 109 atDNA microsatellite loci[note 11] | Barbujani et al. (1997)[46] | 84.4% | 4.7% | 10.8% |
| 79 RFLP's[note 12] and 30 atDNA microsatellite loci | Jorde et al. (2000)[56] | 84.8% | 1.6% | 13.6% |
| 21 Alu insertions | Romualdi et al. (2002)[57] | 82.9% | 8.2% | 8.9% |
| 377 atDNA microsatellite loci[note 11] | Rosenberg et al. (2002)[58] | 94.1% | 2.4% | 3.6% |
| 100 Alu insertions | Watkins et al. (2003)[59] | 88.6% | 1.9% | 9.6% |
| 17 X chromosome markers | Ramachandran et al. (2004)[60] | 90.4% | 4.6% | 4.9% |
| 40 atDNA insertions/deletions | Bastos-Rodrigues et al. (2006)[61] | 85.7% | 2.3% | 12.1% |
| 642,690 atDNA SNPs[note 13] | Li et al. (2008)[62] | 89.9% | 2.1% | 9.0% |
| 783 atDNA microsatellite loci[note 11] | Rosenberg (2011)[63] | 93.9% | 2.4% | 3.8% |
| 27 landmarks and dimensions of the pelvis | Betti et al. (2013)[64] | 91.5% | 5.6% | 3.0% |
This is not to say that there is no variation between humans. As the above table should make clear, it's quite the opposite. Indeed, genetic data does show that random people sampled from within a population are generally more similar to each other than a pair sampled from different populations.[35] (That's hardly surprising since, when left to their own devices, people tend to procreate with local folk more than with folk from another part of the world.) Moreover, with enough genetic information (i.e. more than 1000 genetic loci), it is possible to quite accurately identify the geographic origin of a sampled person's ancestors, but only when the chosen regions are quite distinct.[35] (Or: Icelanders and Ashkenazi Jews might be genetic clusters, but "Caucasians", "Mongoloids", and "Negroes" are not). However, Witherspoon 2007 cautions:[35]
The fact that, given enough genetic data, individuals can be correctly assigned to their populations of origin is compatible with the observation that most human genetic variation is found within populations, not between them. It is also compatible with our finding that, even when the most distinct populations are considered and hundreds of loci are used, individuals are frequently more similar to members of other populations than to members of their own population. Thus, caution should be used when using geographic or genetic ancestry to make inferences about individual phenotypes.
In other words: Just because variation exists does not mean it is meaningful. As the above sections about race in animals should make clear, a little variation does not a race make.
Theoretical and practical flaws
Definitional issues
“”[W]hile some minimal revision to the meaning of 'race' [...] is allowable in the search for biological backing for race, we must stay fairly close to the vest, or we risk not talking about race at all. |
| —Glasgow 2003[65] |
The main problem is that new concepts of race used by racialists are watered down to the extent they are trivialised. Hochman 2014 writes: [66]
The problem with weak versions of racial naturalism is that they do not contrast with anti-realism about biological race. When race naturalists weaken their position they end up agreeing with their opponents about human biology, and defending a trivialised definition of race.
Hochman (2013) rejects these new concepts and re-definitions on the grounds (emphasis added) "the criteria applied to humans are not consistent with those used to define subspecies in nonhuman animals, and no rationale has been given for this differential treatment".[67]
Another problem with re-defining race is: "To avoid making 'race' the equivalent of a local population, minimal thresholds of differentiation are imposed".[68] Modern racialists however argue there is no threshold which runs into the problem of any population being a race: "There are undoubtedly no two genetically identical populations in the world; this has nothing to do directly with the validity of race as a taxonomic device. Unless we have defined exactly what we mean by this… differences between populations are population differences, nothing more."[69] Another objection is a "mismatch argument", where local breeding populations not ordinarily conceived as races (e.g. Amish, or Kalash people
Epigenetics
Along the way from chemistry to expression, the environment gets its chance to intervene. Epigenetics studies the way in which factors outside of the DNA itself alter the way in which genes are expressed. Resembling Lamarckian genetics, epigenetic effects can be remarkable enough for a mouse to pass a specific fear response down at least two generations (F3).[71] In humans, increased risk for cardiovascular disease among African Americans is hypothesized to result from genetic expression modified by in utero experience of maternal stressors.[72] Epigenetic effects might tie socioeconomics to future IQ, explaining some portion of controversially observed racial differences.[73][note 14]
Case study: Caucasoid, Mongoloid, Negroid
As an example of why arguments for racialism fail, consider the case of the "-oid" concept of race.
Ethnic groups and demes in Africa/East Asia/West Eurasia do not form a sharply discontinuous genetic (or phenotypic) cluster, nor are there substantial differences between populations in East Asia, compared to populations from other regions across the globe. Overall, genetic variation between continental human populations is fairly little (<10%) and does not support the concept of different human races. The terms, however, have continued into modern forensic literature (though they are being phased out even of that).
For several reasons, these types of theories are incredibly flawed.
Capoids and Australoids
Carleton S. Coon
Coon did not, however, figure out what "oid" the Native Americans were. Perhaps racialists of the future will finally create the long-anticipated 6-oid system — or find out that Native Americans were actually just really tanned Caucasoids.
Reversed hierarchy
Funnily enough, the cranial order was once asserted to be Caucasoid > Mongoloid > Negroid; clearly, whites are the bestest. Nowadays, as racialists have become fond of Asians, the order has shifted to Mongoloid > Caucasoid > Negroid. Perhaps an entire continent of people have simply upgraded their skulls?[74]
Alternate classifications
Why stop with simplistic appearance-based racial definitions? Why not consider other definitions for which there are genetic determinants?
- Type B blood: Native Americans (First Nations for residents of Canuckistan) and Aboriginal Australians would form one race while Africans would form up partly with Europeans and partly with Asians.[75]
- Sickle-cell anemia risk: One race would encompass the major malaria-risk zones of Central and Northern Africa, parts of the Mediterranean, Arabia, and India.[76]
- Lactase persistence, the ability to digest milk sugar in adulthood: much more common in areas where dairy farming has been practised for thousands of years.[77]
- Defined by one gene alone, you could divide the world into sprinters (strong RR on ACTN3), long-distance runners (strong RX on ACTN3), and everyone else.[42][43] Geographically, that's some people with West African heritage, some with East African Heritage, and everyone else.
Race and intelligence
One obsolete theory attempted to link cranial capacity (cc) to intelligence. While cranial capacity is an accurate measure of brain size, the fallacy was in assuming that a larger cranial capacity (and brain) correlates with higher intelligence. The size of the brain itself is only weakly correlated with intelligence.[78]
If this idea were taken to its logical conclusion, the world would likely be ruled by elephants, or sperm whales. It is only when a person has an extremely small brain, as with abnormal conditions such as microcephaly,
Furthermore, having a very large amount of brain tissue, referred to as megalencephaly, is recognized as being pathological and is strongly correlated with several neurological disorders, especially severe epilepsy and autism. What is much more important than absolute brain size includes factors such as cortical folding, neuronal organization, dendritic and synaptic connections, etc.[79]
A handful of 21st century racialists including figures such as J. Philippe Rushton and Arthur Jensen have continued to argue that certain races are just inherently dumb. While they still like their skull and brain size measurements, their arguments hinge more on pointing to differences in races' average IQ scores and claiming this is the work of genetics. These claims rest on several big assumptions:
- That IQ is a measure of some kind of objective intelligence, rather than a measure of the skills needed to excel in 21st century Western society (a controversial claim).
- That there are genetic differences between "races" big enough to explain the IQ difference.
- That IQ is more dependent on racial genes than environment (or: if environment affects IQ, then the differences in IQ by race should still be significant after controlling for the environment).
Increasingly, evidence has been suggesting that environment plays a large role in IQ. This started with the discovery of the Flynn effect — the realization that national average IQ scores were increasing over time at a rate much faster than could be explained by genetics (and, interestingly, ethnic minorities were often making the biggest leaps).[80][81]
Relevant to this discussion also is Eric Turkheimer's study of twins separated at birth and raised in different socioeconomic environments. The study found that environment mattered more to the development of IQ in the poor kids than it did in the rich kids. Essentially, some degree of genetically-determined IQ exists, but the degree to which it develops is dependent on how intellectually-stimulating their environment as children is.[73] Other explanations, such as the stereotype threat, racial discrimination in the education system, lack of funding for schools in poor areas, more talented teachers avoiding teaching at schools in poor areas, and even differences in diet have been offered.
An irony
Although these IQ-based arguments for racialism are often used by white supremacists, taken to their logical conclusion, they would better support Asian or Jewish supremacy than white supremacy. Rushton actually agrees that Asians, not whites, are the intellectual master race, leading anthropologist Jonathan Marks to remark:[74]
First we must admire the apparent cranial expansion of Asians over the last half-century, when [earlier] researchers consistently reported their having smaller brains than whites. Obviously this implies the possibility of a comparable expansion in blacks. More likely, it implies the possibility of scientists finding just what they expect when the social and political stakes are high.
Adaptation doesn't work like that
Adaptation to environments, including social environments, through natural and sexual selection of random mutations is the linchpin of evolution. Remembering this means knowing why scientific racism is ridiculous. To argue that races or ethnic groups differ innately in intelligence, however defined, is exactly equal to an assertion that intelligence has proven less adaptive for some people than for others.
This at minimum requires an explanation, a specifically evolutionary explanation, beyond mere statistical assertion; without that it can be assumed to be cultural bias or noise. Since most human intelligence is in fact social intelligence — the main thing the human mind is built for is networking in human societies — this would require this social evolutionary arms race to have somehow stopped.
One such explanation could be that within a certain population the random mutations needed for certain intelligence boosts never occurred. The genetics related to brain development are not uniform among the genetically diverse and geographically distant populations (the microcephalin alleles being an example of brain-related gene complexes that are not geographically uniform).[82] Wade 2005 writes:[82]
They report that with microcephalin, a new allele arose about 37,000 years ago, although it could have appeared as early as 60,000 or as late as 14,000 years ago. Some 70 percent or more of people in most European and East Asian populations carry this allele of the gene, as do 100 percent of those in three South American Indian populations, but the allele is much rarer in most sub-Saharan Africans. With the other gene, ASPM, a new allele emerged some time between 14,100 and 500 years ago, the researchers favoring a mid-way date of 5,800 years. The allele has attained a frequency of about 50 percent in populations of the Middle East and Europe, is less common in East Asia, and found at low frequency in some sub-Saharan Africa peoples.
To argue otherwise is to argue that all required mutations magically appear in all social living beings as soon as they develop social intelligence.
Genetic differences between people from different locations are pretty much entirely genetic drift (which itself can lead to the appearance of genes that are not expressed anywhere else). Amount of melanin does vary with distance from the equator, and there's recent actual evolution such as Lactase persistence in adults.[83]
Race and sporting ability
There are several common beliefs about how different races have different sporting abilities. Clichés include:
- Blacks are good at track and field (especially sprinting[84]) and heavyweight boxing (hence the phrase "The Great White Hope"[85]).
- Blacks can't swim.[86]
- East Africans are good at long-distance running.[87]
- East Asians are athletically weaker, at least at some sports.[88][89]
- Whites are better at tennis.[90] (except the Williams sisters
File:Wikipedia's W.svg ). - Whites can't jump (basketball).[91]
On the other hand, why has Norway (population 5 million) won 132 golds in Winter Olympics and Vietnam and India none?[92] Are Asians intrinsically terrible at skiing compared to tall Nordic types? Or does Norway have more ski slopes and better ice rinks and just care more and spend more money on it? Why has one small Catholic University
It appears that some factors implicated in sporting success are heritable (e.g. height), and therefore will be more or less common in groups with common ancestors. However, in many cases, these do not correlate closely to race (the Dutch are the tallest nationality[93], despite stereotypes about black basketballers). As the section on genetic variation makes clear, there is not a simple pot of genes that corresponds to each race, so you can't assume a member of a race will have a given gene. And there are few clear cases where genes define sporting success: even factors like height are affected by nutrition, medical facilities, etc, and distributed across races.
One well-known genetic factor is the gene ACTN3 associated with fast twitch muscle
It is also important not to confuse genetic heritage with nationality, cultural factors, or social status. There are many non-genetic reasons why people of a certain race, ethnicity, or nationality may choose to play specific sports, or excel at them. These include:
- economic factors (which may affect everything from nutrition and health, to ability to afford equipment and tuition, and time to train)
- discrimination (from racist sports clubs restricting membership, to expectations of coaches)
- environment and climate (where you live may affect what sports you do, along with factors such as heat or cold tolerance or lung capacity developed from living at high altitude)
- religions or cultural factors (e.g. requirements that women keep their bodies covered, or stay in the home)
- cultural beliefs about the importance of specific sports or sport in general (anecdotally some cultures celebrate sport as a means to success, while others favour academic achievement at school)
- Cultural ties of a sport to a certain ethnic group. For instance, within South Africa, Rugby Union was long considered the sport of white Afrikaans speaking people while cricket was considered the sport of white people of English descent with soccer the sport of everybody else. This has only begun to change since Apartheid ended.
Sport science academic Ben Oakley attributes the success of east African long-distance runners partly to their experience at high altitude and partly to the cultural factors that encourage running in east Africa more than in high regions of Latin America. According to Oakley there is no specific genetic factor, but their tendency to light, lean bodies probably helps. None of these are unique to the region, but they combine to produce a greater effect.[87]
Race and medicine
A topic that has been controversial (for both social and scientific reasons) is so-called "race-based medicine." Currently, some doctors may use racial profiling as a proxy for determining the geographic origins of the patient's lineage to diagnose and treat certain diseases.[96] However, this is not the same thing as "race." The researchers at the Human Genome Project sum it up:[97]
DNA studies do not indicate that separate classifiable subspecies (races) exist within modern humans. While different genes for physical traits such as skin and hair color can be identified between individuals, no consistent patterns of genes across the human genome exist to distinguish one race from another. There also is no genetic basis for divisions of human ethnicity. People who have lived in the same geographic region for many generations may have some alleles in common, but no allele will be found in all members of one population and in no members of any other.
The word cline is sometimes used to describe this. To put it more succinctly, as the leader of the Human Genome Project, Francis Collins does:[98]
“”'Race' and 'ethnicity' are poorly defined terms that serve as flawed surrogates for multiple environmental and genetic factors in disease causation, including ancestral geographic origins, socioeconomic status, education and access to health care. |
Clarification: Heuristics versus biology
It should be noted that all human classifications used in medicine are considered probabilistic heuristics and not perfectly deterministic. While research has shown that many people are extremely effective at classifying themselves based on ethnic origin[99] and so researchers may find "race" to be a convenient label, it fails as a source of attribution: "[R]ace as a social construct may result in differences in treatment that affect health outcomes, but such descriptive use does not imply that "race" can be used as a proxy for biological difference."[100]
"Race" will continue to exist as a cultural phenomenon; while great care should be taken in using this construct in medicine, it would be counter-productive to discard potentially useful information, such as the way in which racial identity shapes social, legal and economic outcomes. Unfortunately, the knotty history of "race" makes even the language used a potential source of error — "race realists" often point to a news headline or study that uses "race" sloppily and declare it to be "evidence" of the truth of "race," mastering sufficient jargon to talk about the reality of "genetic clines" and missing the fundamental mismatch between "race" as culture and "race" as genetic destiny.
Current trends: Against racialism
Even if race was useful or currently is useful, current medicine is moving towards actually looking at people's genetics (rather than their phenotype (race), which is a poor approximation). As Pena 2011 puts it:[101]
Personalized pharmacogenomics aims to use individual genotypes to direct medical treatment. Unfortunately, the loci relevant for the pharmacokinetics and especially the pharmacodynamics of most drugs are still unknown. Moreover, we still do not understand the role that individual genotypes play in modulating the pathogenesis, the clinical course and the susceptibility to drugs of human diseases which, although appearing homogeneous on the surface, may vary from patient to patient. To try to deal with this situation, it has been proposed to use interpopulational variability as a reference for drug development and prescription, leading to the development of “race-targeted drugs”. Given the present limitations of genomic knowledge and of the tools needed to fully implement it today, some investigators have proposed to use racial criteria as a palliative measure until personalized pharmacogenomics is fully developed. This was the rationale for the FDA approval of BiDil for treatment of heart failure in African Americans. I will evaluate the efficacy and safety of racial pharmacogenomics here and conclude that it fails on both counts. Next I shall review the perspectives and the predicted rate of development of clinical genomic studies. The conclusion is that “next-generation” genomic sequencing is advancing at a tremendous rate and that true personalized pharmacogenomics, based on individual genotyping, should soon become a clinical reality.
Case study: Sickle-cell anemia
Sickle-cell anemia is a famous example of how "race" can mislead. It was originally classified as a "negro disease" due to its high incidence in blacks. However, it was later found that sickle-cell anemia, being an adaptation to the risk of malaria, was also more common in central and western Africans (but not southern Africans) as well as Mediterraneans (Turks, Greeks, etc.) and Indians. Thus, the disease was not an indication of "race," but of geographic ancestry (in this case, areas where malaria was more common).[102]
Race as disease risk categories?
Classification of race in medical genetics (e.g. "Caucasian", "Black") has been criticised on the grounds studies on disease rarely control environmental factors and so a genetic etiology is assumed without testing. That said, no doctor denies that some diseases and found at high frequency in some populations, while low in others and therefore categorisation may provide useful. While information about ethnicity may be informative for biomedical research: "it is imperative to move away from describing populations according to racial classifications such as 'black', 'white' or 'Asian'… there can be considerable genetic heterogeneity within a region, it is most useful to be as specific as possible about geographic origins, ethnicity or tribal affiliation".[103] Those diseases that show considerable inter-group difference, are only confined to local breeding populations, that are often more inbred (e.g. ethno-religious sects such as the Samaritans) rather than large regions, or continents. Dr. Jurgen K. Naggert, a geneticist at the Jackson Laboratory in Bar Harbor, Maine, has noted:[104]
“”These big groups that we characterize as races are too heterogeneous to lump together in a scientific way. If you're doing a DNA study to look for markers for a particular disease, you can't use 'Caucasians' as a group. They're too diverse. |
ApoL1 associated kidney diseases are a lot more common in many populations in Africa than Europe and Asia, to the extent they've been incorrectly described as a 'black disease', but because Africa is a massive continent there are some countries (e.g. Ethiopia) where ApoL1 associated kidney diseases are in fact absent: "ApoL1 risk variants nor the sickle-cell allele are 'black' diseases because many 'black' populations do not have these alleles.[105] Another example, α-thalassemia has a high frequency (80%) in an ethnic group in South Asia (Tharu people in Nepal), but very low in others, so it would be meaningless to talk of an 'Asian disease' (Lewontin, 2006).[106]
Race and forensic anthropology

Racialists quote forensic anthropologists who are said to be able to estimate ancestry of skeletal remains by continent ("race") with an accuracy of up to 90%. For example, a 1986 textbook by forensic anthropologists claimed that race should be determinable from skull morphology alone with an accuracy of 85-90%.[108]:296[109] The revised version of the textbook did not make such a claim however, and cited Hefner (2009)[110] that "'the lack of a methodological approach' used in these methods, and the fact that there are no error rates associated with ancestry prediction using them. He suggested that they have not been investigated with appropriate scientific and legal considerations in mind, and aspects such as inter-observer repeatability have hardly been considered. … forensic anthropologists claim that they can accurately asses ancestry using these [skeletal] traits, and the actual frequencies of these traits are, in fact, much lower than assumed. … [Ousley (2006)] suggested that analysts make a diagnosis of ancestry based on an overall impression of a skull, and then choose the traits post-hoc to support their assumption as a type of confirmation bias."[111]:203
Empirical flaws: Sampling bias
The high accuracy figures quoted above only apply to limited subsets of a sample. However, this high accuracy figure only matches limited subsets of a sample in each broad continental grouping, and "in cases where independent samples are used to test the methods, allocation accuracies decrease to levels that undermine the applicability of the methods in actual cases."[112] As such, broad groupings like "Mongoloid" have little to no utility.[113]
Two case studies should drive home this point. First consider Birkby 1966, who found a high allocation accuracy for "Native Americans" (>90%) based on an archaeological sample from Knoll, Kentucky.[114] When however indigenous remains were sampled from many other locations, the accuracy for the "Native American" racial category diminished: "the methods performed poorly on the respective American Indian samples (accuracies ranged from 14 to 30%), and confirmed Birkby’s conclusion that the Indian Knoll sample cannot be considered a proxy for the pattern of variation in numerous populations that are included in the group American Indian".[115]
Similarly, Smay & Armelagos 2000 write:[112]
[T]he 85 percent to 90 percent results are based on the accuracy of the method when standards are developed on a subset of a sample and then "tested" on the sample from which the subset was derived. For example, in the 1960s when Eugene Giles and Orville S. Elliot (1962) developed their formula to determine race from the crania, they used a sample that was a sub-set of modern adult Blacks, modern adult Whites and Native American skulls from an archeological site (Goodman 1997b). They applied a statistical procedure—discriminant function that separates crania into “races” using eight measurements. When they applied the formula to the rest of the crania in the same sample, they achieved the much touted 85 percent to 90 percent accuracy. When applied to other samples of Blacks, Whites and Native Americans, they achieved 18.2 percent and 14.3 percent accuracy, figures that hardly instill confidence (Goodman 1997b).
Empirical flaws: Inconsistent methods
The high accuracy rate claim for racial determination methods is also questionable since different methods have a tendency to produce different results:[116]
We have shown that even with 20 non-fragmented sets of skeletal remains none could be consistently placed into a single racial category. Individual variability may have played a significant role leading to inconsistency of the results found in this study, which further confirms the ideas of Brace and Ryan (1980), Henneberg (2010) and Lewontin (1976); that most human variation occurs between individuals of the same population rather than being attributable to geographic distribution.
Empirical flaws: Gradients of features within races
The fact that there are gradients of features within so-called races is widely recognized and contradicts discrete racial boundaries. As an example, let us consider Asia. According to Wu Rukang and Wu Xinzhi (1997): "The cephalic, facial, and nasal indices, plus stature, show clinal variation. From north to south [China], the cephalic index and the stature are seen to decrease, while the facial and nasal indices increase."[117]
This can easily be seen with an example. Ling & Wong (2008) provide a table of shoveling among East Asian populations, with results for tooth I1 reproduced below:[118]
| Ethnic Group | Gender | Shovel (%) | Semi-Shovel (%) | Trace (%) | None (%) |
|---|---|---|---|---|---|
| Southern Chinese | M | 7.0 | 66.0 | 27.0 | 1.0 |
| F | 3.0 | 68.0 | 27.0 | 1.0 | |
| Taipei Chinese | M | 75.9 | 20.1 | 4.0 | |
| F | 85.3 | 13.2 | 1.5 | ||
| Chinese | M | 66.2 | 23.4 | 1.8 | 7.8 |
| F | 82.7 | 12.5 | 1.0 | 3.8 | |
| Japanese | M | 76.1 | 15.1 | 3.1 | 2.3 |
| F | 74.1 | 16.2 | 2.7 | 1.1 | |
| Liverpool Chinese | 77.5 | 21.7 | 0.8 | 0 | |
| Eskimos | 37.5 | 47.5 | 15.0 | 0 | |
| Mongolians | 62.5 | 29.0 | 8.5 | ||
| Aleuts | M | 64.4 | 31.1 | 4.5 | 45 |
| F | 60.0 | 40.0 | 30 | ||
Full shoveling spans from 85.3% to 3.0% among Chinese populations alone.
Theoretical flaws: Region versus race
Sauer (1992) notes that "to estimate, with varying degrees of specificity, a person’s place of ancestry from their physical features" is "not a vindication of the [biological] race concept".[109]
C. Loring Brace makes the same point that "region, does not mean race".[119] More recently terms such as "geographical ancestry", or "ancestral population" have replaced race in forensic literature.
Motivation of forensic anthropologists
If race is so meaningless, why do so many forensic anthropologists still use it? The simple answer is that laypeople often require them to do so. Smay & Armelagos 2000 write:[112]
Many forensic anthropologists understand race to be a folk taxonomy with little, if any, biological relevance. Given this, however, they point to the duty of forensic anthropologists to serve the medico-legal communities to which they have an obligation. These communities are not interested in the fact that race does not exist, and, according to Sauer and Kennedy, are not likely to be convinced of this in the near future. On the contrary, race is used as a key element in the search for missing persons, and forensic anthropologists are expected to provide this information in their reports. Even if races are not biologically "real", the accuracy with which forensic anthropological techniques are able to replicate the folk taxonomy from which they are derived should allow the anthropologist to make an educated guess as to how the person would have been identified in life.
"Lewontin's fallacy"
Most modern racialists redefine race as a "genetic cluster" by continent which captures a fairly small amount of variation (<10%) between groups of populations. Molecular anthropologists such as Jonathan Marks are confused by this re-definition because it is so far removed from biological taxonomy, and the traditional meaning of race. Michael P. Muehlenbein writes:[120]
What is unclear is what this has to do with 'race' as that term has been used through much in the twentieth century — the mere fact that we can find groups to be different and can reliably allot people to them is trivial. Again, the point of the theory of race was to discover large clusters of people that are principally homogeneous within and heterogeneous between, contrasting groups.
The geographical pattern to human interpopulation variation, matches an almost continuous gradient/isolation-by-distance model[121][122][123][124][125][37] However much of this discontinuity is found within continents, not between them[46][126][127] — which again renders the ideas of (e.g.) "white" and "black" as irrelevant.
Racialists argue that while overall genetic variation between continental population divisions is <10%, this rather little human population structure still supports race classification (contra Lewontin, 1972[51]).
They quote Edwards (2003) who found by looking at how gene loci are correlated: "probability of misclassification falls off as the number of gene loci increases". It is notable Edwards in his paper does not dispute Lewontin's statistical data on various blood polymorphisms, writing: "There is nothing wrong with Lewontin's statistical analysis of variation, only with the belief that it is relevant to classification". As a result, as Marks (2010) explains, there is no "Lewontin fallacy":[128]
“”Geographical correlations are far weaker hypotheses than genetically discrete races, and they obviously exist in the human species (whether studied somatically or genetically). |
What Lewontin (or Marks) and Edwards are discussing are two completely different things. Of course genetic correlations exist which can pinpoint someone's geographical ancestry, but as Marks asks: "What is unclear is what this has to do with race", and concludes: "Lewontin's analysis shows that such groups [races] do not exist in the human species, and Edwards's critique does not contradict that interpretation" (emphasis added). What this means is that Edwards is re-defining the race concept to a far weaker hypothesis, which is not how race is commonly understood in biological taxonomy: Fujimura 2014 writes:[123]
“”[I]f one wants to measure the meaningfulness of differences between groups with different geographic ancestries, one needs to use a “proportion of variation” approach. This approach compares genetic variation among individuals within groups to the genetic variation between these groups. Ironically, Edwards’s (2003) reported findings confirm Lewontin’s (1972). |
What is the significance of the "proportion of variation" (FST) approach? It measures phylogenetic 'treeness' via genetic differentiation[47] i.e. whether populations reflect a common evolutionary history or intraspecific distinctive lineage, which is taxonomically meaningful. Human populations however "have such little structure that ‘treeness’ is not demonstrated and phylogenetic models are invalid".[17] As Lewontin 1974 himself writes:[129]
The taxonomic division of the human species into races places a completely disproportionate emphasis on a very small fraction of the total of human diversity. That scientists as well as nonscientists nevertheless continue to emphasize these genetically minor differences and find new “scientific” justifications for doing so is an indication of the power of socioeconomically based ideology over the supposed objectivity of knowledge.
FST
FST (or the fixation index

Wright (1978) calculated FST values under 5% indicate little genetic differentiation between populations, 5% to 15%, moderate genetic differentiation, 15% to 25%, great genetic differentiation, and above 25%, very great differentiation which is the threshold for subspecies (races).[67] Human continental population divisions fall at the lower end of moderate genetic differentiation (<10%), while demes and ethnic groups under 5%: "Hence, as judged by the criterion in the nonhuman literature, the human FST value is too small to have taxonomic significance under the traditional [definition of] subspecies".[68]
Muddying the waters
In order to discredit the science against racialism, it is a common racialist tactic to compare race with some other (supposed) biological characteristic. For example, as one commenter on The Right Stuff writes:[130]
Lewontin's Fallacy, that because there is more variation within a group than between groups, therefore the groups are the same.
The difference in average height between men and women is about 4" (7%), but the variation in height among women is about 24" (4' to 6') , and among men about 36" (4' to 7'), therefore men and women are the same average height. Clearly false.
The commenter fails to note that human sexes are (traditionally) differentiated on the grounds of certain attributes which have decidedly low rates of variation within a sex. (Or: the genes considered "female" almost always result in an organ-level difference from those for "male".) In other words: the variation in height is not what maleness and femaleness are based on, and so variation within them doesn't matter. In contrast, the variation in certain traits (eg, skin color) are precisely what race is based upon, and so variation within one "race" being greater than between races makes it functionally meaningless.
Identification of race via genetics
Similar to creationist quote-mines of evolutionist literature, online racialists often do the same with Witherspoon et al. 2007:[35]
Thus the answer to the question “How often is a pair of individuals from one population genetically more dissimilar than two individuals chosen from two different populations?” depends on the number of polymorphisms used to define that dissimilarity and the populations being compared. The answer, Formula can be read from Figure 2. Given 10 loci, three distinct populations, and the full spectrum of polymorphisms (Figure 2E), the answer is Formula ≅ 0.3, or nearly one-third of the time. With 100 loci, the answer is ∼20% of the time and even using 1000 loci, Formula ≅ 10%. However, if genetic similarity is measured over many thousands of loci, the answer becomes “never” when individuals are sampled from geographically separated populations.
The above paragraph is what they spam, especially the bold. What they don't post is the paragraph that immediately follows:
On the other hand, if the entire world population were analyzed, the inclusion of many closely related and admixed populations would increase. This is illustrated by the fact that Formula and the classification error rates, CC and CT, all remain greater than zero when such populations are analyzed, despite the use of >10,000 polymorphisms (Table 1, microarray data set; Figure 2D).
Two individuals from separate populations can then be genetically more similar than two individuals from the same population (with >10,000 SNP's). In other words: even with an incredibly stringent set of criteria (think about the last time you sat down and listed out 10,000 genetic reasons that someone was a member of their race), race still isn't a binary, but a gradient. Moreover, at the point where you are using >10,000 polymorphisms, races like "white" and "black" have utterly ceased to exist and are replaced with innumerable smaller groups.[note 15]
Claims of bias in anthropology
“”The rejection of the race concept […] in the 1960s was based on the genetic evidence reviewed earlier. Conformity to political correctness was not the cause of these changes[.] |
| —Lieberman et al 2003[2] |
What now?
“”Race as biology is fiction, racism as a social problem is real |
| —Smedley and Smedley 2005[131] |
Race isn't biologically useful. But it still carries substantial social importance that we can't just wish away. For now, the best option may be to educate people about the insignificance of race and (as racial parity is achieved) hope race fades from social significance. As Cartmill 1998 writes:[33]
Like other social constructs, races are real cultural entities. For many people, membership in a racial group constitutes an important part of their social identity and self-image. But social facts are not necessarily part of the biological landscape. In multiethnic regional populations, races are merely ethnic groups linked to vague, inconsistent, and stereotypical ideal phenotypes. Growing awareness of the meaninglessness of racial taxonomy is currently leading increasing numbers of U.S. citizens to refuse to classify themselves racially, or to allow themselves to be so classified by others (Fish 1995). In the long run, we would probably be better off if we all followed their example.
Notable racialists
- Benjamin Rush: Rush was, rather interestingly, one of the Founding Fathers and an abolitionist. He believed that blacks suffered from a disease called "negritude," supposedly a form of leprosy, that could be "cured" and would result in turning their skin white.[132] This led to his attempts to develop quack "cures" for "negritude."
- Samuel Cartwright:
File:Wikipedia's W.svg Expanding on Rush's work, Cartwright argued that drapetomania also a physical illness that could be diagnosed through the appearance of lesions on the skin. No doubt this had nothing to do with the backbreaking manual labor and frequent whippings and beatings slaves endured.[132] Cartwright coined two pseudo-psychological diagnoses that rationalized and justified slavery. The first he called "drapetomania," which was allegedly an illness causing slaves to run away from their masters. Conveniently for the slave masters, much of the "treatment" for this "illness" consisted of whipping. The second "diagnosis" was "dysaethesia aethiopica", or laziness.
- J. Philippe Rushton: Advocate of "Life History Theory" that proposes that all socially desirable personality and intellectual traits are concentrated in the white and Asian races, whereas all the antisocial and undesirable traits are concentrated in black people. Repeatedly criticised in academia for sloppy scholarship including using questionable sources of information including the Penthouse Forum.[133]
- Richard Lynn: Eugenics advocate who recently wrote a paper claiming that black people have longer penises than whites, who in turn are better endowed than Asians.[134] This was based on Rushton's "Goldilocks" theory of race: "Orientals have big brains but small genitalia, Africans have small brains but big genitalia, but Europeans Are Just Right."[135] One of the key sources of data used in this paper was the World Penis Size Site,[136] an anonymously compiled source that includes a lot of made-up data and bogus references.[137] Lynn has also stated that "incompetent cultures" need "phasing out."[138] Just to clear up any doubt about which cultures he thinks need "phasing out" he writes: "Who can doubt that the Caucasoids and the Mongoloids are the only two races that have made any significant contributions to civilization?"[139]
- Donald Templer: Authored a book on penises used as an additional reference by Richard Lynn even though Templer has no qualifications in urology. Advocates the voluntary sterilization of welfare recipients on eugenics grounds.[140][141]
- Lothrop Stoddard:
File:Wikipedia's W.svg : American racist, eugenicist, and anti-immigration campaigner, author of many books including The Rising Tide of Color Against White World-Supremacy. He was enormously influential in early 20th century America, and is satirised by F. Scott Fitzgerald in The Great Gatsby, but a lengthy and enthusiastic visit to Nazi Germany in 1939-40 caused his reputation to fall after America entered World War II.
- Varg Vikernes: Norwegian musician, arsonist, murderer, gun lover, authoritarian wingnut, Neopagan, Neo-Nazi, White nationalist, anti-Semitic conspiracy theorist, anti-vaxxer, prepper and former YouTube personality, banned recently from the platform.
- David Duke: former Grand Wizard (what?) of the Ku Klux Klan (yes, really).
- Stefan Molyneux: former YouTube personality, banned recently from the platform
- Jared Taylor: editor of American Renaissance, a magazine for white nationalism that he founded
- Richard Spencer
- Sargon of Akkad
- Nazis
- Metapedia
- Human Phenotypes
- Human Varieties
External links
- Academic sources:
- American Anthropological Association's statement on "race"
- Is Race "Real"? a forum by the Social Science Research Council
- Institute for the Study of Academic Racism
- Anthropology and Race, Anthropology Report
- Biological Anthropology: Human Nature, Race, Evolution
- Biological Races in Humans
- Popular sources:
- IQ and Race (The Skeptic's Dictionary)
- Biologically Speaking, Race Doesn't Exist, Fox News (2002)
- THERE IS NO SUCH THING AS RACE, Newsweek (2014)
- Classifying humans into races the biggest mistake in the history of science, ABC (2016)
- Videos:
- Is race a social invention?: A video discussion by a biological anthropologist going into the problems with attempting to define race biologically
- What Can DNA Really Tell Us About Race?, Troy Duster, UCtelevision
- Race, the human spectrum
Further reading
- Edgar, Heather and Keith Hunley. "Race reconciled?: How biological anthropologists view human variation". American Journal of Physical Anthropology, May 2009, Volume 139, Issue 1, Pages 1–4.[1]
- Lieberman, Leonard, et al. "The decline of race in American physical anthropology". Przegląd Antropologiczny – Anthropological Review. 2003, Vol. 66, 3-21.[2]
- Genetically Speaking, Race Doesn't Exist In Humans, EurekAlert (1998)
- Lynn B. Jorde and Stephen P. Wooding. Genetic variation, classification and 'race'. Nature Genetics 36, S28 - S33 (2004)
- Kenneth M. Weiss and Stephanie M. Fullerton. Racing Around, Getting Nowhere. Evolutionary Anthropology 14:165–169 (2005)
- Gurdasani, Deepti, et al. "The African genome variation project shapes medical genetics in Africa." Nature 517.7534 (2015): 327-332.
Notes
- While scientific racists do have to admit their racism, they also get the joy of slapping the "approval" of SCIENCE on it.
- Racial realists get a two-pronged advantage: First, they can deny their racism by saying, "I'm not a racist, I'm a realist!". Second, they can paint non-racialists as "race deniers" or "racial difference deniers", which suggests denial of the obvious facts — when in fact, science opposes racialism.
- By being just one "bio" away from supporting "diversity" and by failing to mention race, human biodiversity is the most innocuous form of racialism, because it allows one to deny any racism — because it's recognizing biodiversity, not being racist! Coined by Steve Sailer (a political blogger) of VDARE. Unlike biological diversity, HBD is unscientific.
- However, there was substantially more disagreement on the role of race in medicine and race in social issues, both of which are discussed later in the article. As Wagner write:
While a closer examination of perspectives of biological anthropologists not affiliated with AAA or attending AAA meetings is needed, our data (including those shown in Table 6) suggest biological anthropologists and particularly those with familiarity with genetic ancestry testing might be more willing to acknowledge that race—not understood as a biologically-defined category but, rather, as a socially-defined category and as a lived experience of structural and institutional racism—can have important effects on health.
- The TLDR is available here.
- The correlation is the Spearman's rank correlation coefficient
File:Wikipedia's W.svg , rho. Positive correlation means that familiarity with genetic ancestry inference and proportion of participants with the common response tended to both increase at the same time (or: response and familiarity are more monotonous). TLDR: Positive means familiarity and the common response went together; negative means the opposite. The p-value is in parentheses. Asterisk indicates significance. - Y-chromosomes are passed down through fathers. For the female equivalent (passed through mitochondria), see Human mitochondrial DNA haplogroup
File:Wikipedia's W.svg . - 17 blood groups.
- 10 blood groups, 3 serum proteins and 5 red blood cell enzymes.
- 14 serum proteins, 9 blood groups and 2 leukocyte antigens.
- Autosomal DNA (meaning, any DNA inherited from chromosomes other than sex chromosomes) microsatellite (meaning, long stretches of DNA where a pattern in the DNA is repeated) locations.
- Restriction fragment length polymorphisms (meaning, variations in the length of a restriction fragment, which is a fragment of DNA that has been cleaved by a restriction enzyme).
- Autosomal DNA (meaning, any DNA inherited from chromosomes other than sex chromosomes) single nucleotide polymorphisms (meaning, variations in a single pair of the ATCG nucleotide bases).
- See also the lead-crime hypothesis article.
- Imagine a hooded Klansman saying: "Fuck those Group 1467 bastards. I can't believe that they inherited a statistically significantly different set of polymorphisms."
{kind=link}
References
- Edgar, Heather and Keith Hunley, "Race reconciled?: How biological anthropologists view human variation", American Journal of Physical Anthropology, May 2009, Volume 139, Issue 1, Pages 1–4. DOI: 10.1002/ajpa.20995. Full quote:
The points of agreement in the following articles reflect a shared evolutionary perspective that focuses on describing and interpreting the apportionment of biological variation between individuals both within and among groups (see also Lee et al., 2008). We agreed that:
- There is substantial variation among individuals within populations.
- Some biological variation is apportioned between individuals in different populations and among larger population groupings.
- Patterns of within- and among-group variation have been substantially shaped by culture, language, ecology, and geography.
- Race is not an accurate or productive way to describe human biological variation.
- Human variation research has important social, biomedical, and forensic implications.
- Lieberman, Leonard, et al. "The decline of race in American physical anthropology". Przegląd Antropologiczny – Anthropological Review. 2003, Vol. 66, 3-21. Full quote: TLDR: The reason racialism is challenged is perhaps because of political correctness; the evidence on which it is challenged is purely scientific.
Occasionally a student will suggest that the race concept has been rejected because of political correctness arising from the horrified reaction to the holocaust in World War II Nazi Germany. We must acknowledge that political correctness is a generic aspect of human behavior. Without conformity to group expectations human societies would be anarchies devoid of cultural patterns. An example of the negative consequences of political correctness is seen in the widespread belief that there were superior and inferior races. Colonialism, involving the conquest, exploration and slaughter of millions of people in the Americas, Africa and parts of Asia was rationalized by the belief in the inferiority of the alleged races inhabiting those areas [COCKER 1998]. The rejection of the race concept by most anthropologists beginning most recently in the 1960s, was based on the genetic evidence reviewed earlier. Conformity to political correctness was not the cause of these changes; rather awareness of the uses of race in colonialism, slavery, segregation, and in the holocaust stimulated reexamination of the race concept using the new genetic data that was accumulated throughout the 20th century. The presence of new genetic data does not guarantee that the data will be given careful consideration. This consideration came about as a new generation of anthropologists with new biographical experiences entered the discipline and examined the new data that developed during the century. Those who characterize these developments as political correctness are using simplistic reductionism, and a naive conception of science in an ivory tower. Scientists must struggle with and develop new data in the context of biography and history.
- Wagner, Jennifer K., et al. "Anthropologists' views on race, ancestry, and genetics." American Journal of Physical Anthropology (2016).
- Ruvolo, Maryellen, and Mark Seielstad. "The apportionment of human diversity: 25 years later" in Thinking about Evolution: Historical, Philosophical, and Political Perspectives, edited by Rama S. Singh et al. (2001). Cambridge University Press. ISBN 0521620708.
- Van den Berghe, P. L. (1981). The Ethnic Phenomenon. New York: Elsevier. p. 22.
- Hedrick, P. W. (2011). Genetics of Populations. Jones and Bartlett Publishers. p. 66.
- Templeton, A. R. (2006) Population Genetics and Microevolutionary Theory. John Wiley & Sons. p. 48
- Relethford, J. H. (2012). Human Population Genetics. Wiley-Blackwell. p. 18.
- Templeton, 2013:
“”In traditional taxonomic studies, the boundaries were defined by morphological differences, but now these boundaries are typically defined in terms of genetic differences that can be scored in an objective fashion in all species. Most demes or local populations within a species show some degree of genetic differentiation from other local populations, by having either some unique alleles or at least different frequencies of alleles. If every genetically distinguishable population were elevated to the status of race, then most species would have hundreds to tens of thousands of races, thereby making race nothing more than a synonym for a deme or local population. A race or subspecies requires a degree of genetic differentiation that is well above the level of genetic differences that exist among local populations.
- McKusick, V. A., Bias, W. B., Norum, R. A., & Cross, H. E. (1967). "Blood groups in two Amish demes". Human Genetics, 5(1), 36-41.
- [https://www.encyclopedia.com/social-sciences/encyclopedias-almanacs-transcripts-and-maps/deme Encyclopedia of Race and Racism: Deme.
- Marks, J. M. (1995). Human Biodiversity: Genes, Race and History. New York: Aldine. p. 274:
“”Human populations differ, and often differ from one another on the average in some particular biological characteristics... it comes in lots of small bio-packages
- The Human Genome Diversity Project: past, present and future. Nature Reviews Genetics. 6: 333-340.
- Marks, J. (2010). "Ten Facts about Human Variation" in Muehlenbein, M. P (ed.) Human Evolutionary Biology. Cambridge University Press: 265-276.
- Relethford, J. H. (2017). "Biological Anthropology, Population Genetics, and Race" in The Oxford Handbook of Philosophy and Race. Oxford University Press.
- On the Non-Existence of Human Races by Frank B. Livingstone. Current Anthropology Vol. 3, No. 3 (Jun., 1962), pp. 279-281.
- Deconstructing Race by Caspari, R. (2010). In: Larsen, C. (ed.). A Companion to Biological Anthropology. Wiley Blackwell:
Promoted by Livingstone (1962), the study of clines, the distribution of individual morphological and genetic traits, came to replace race as a focus of analysis for many workers.
- Race or Clines? by Lieberman, L., Rice, P. C. (1996). Arlington, VA: American Anthropological Association.
- Race and global patterns of phenotypic variation by Relethford, John H. American Journal of Physical Anthropology 139.1 (2009): 16-22.
- [web.ornl.gov/sci/techresources/Human_Genome/elsi/minorities.shtml Minorities, Race, and Genomics][a w]
- Biological Aspects of Race[a w]
- AAA Statement on Race[Statement on Race a Statement on Race w]
- Lieberman, Leonard, and Larry T. Reynolds. "The debate over race revisited: An empirical investigation." Phylon (1960-) 39.4 (1978): 333-343.
- Štrkalj, Goran. "The status of the race concept in contemporary biological anthropology: A review", Anthropologist, 2007, vol. 9, no. 1, pp. 73-78.
- Templeton, Alan R. "Biological races in humans." Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, 44.3 (2013): 262-271.
- Smith, H., Chiszar, D., and Montanucci, R. 1997. Subspecies and Classification. Herpetological Review.
- Amadon, Dean. The Seventy-Five Per Cent Rule for Subspecies, 1949. The Condor.
- Remsen, J. V. Jr. SUBSPECIES AS A MEANINGFUL TAXONOMIC RANK IN AVIAN CLASSIFICATION. Onithological Monographs. 2010 no 67 pp 62-78.
- Pasternak, J. (2005). An Introduction to Human Molecular Genetics. Wiley-liss. p. 285.
According to Wright, although the cutoff points are somewhat arbitrary, Fst values that range from 0 to. 0.05 indicate little genetic differentation (genetic dibersity) among subpopulations; 0.05 to 0.15, moderate; 0.15 to 0.25, great; and <0.25 very great... Generally, FST > 0.25 denotes the presence of subspecies within the total population.
- Spencer, Q. (2009). "Is cladistic race a genuine kind?". Stanford University.
Many systematists use a minimum Fst value of 0.25 to mark racial difference.
- Graves, J. L. & Rose, M. R. (2006). "Against Racial Medicine". Patterns of Prejudice. 40(4): 481-493.
Subsequent studies of human genetic variation, including whole genome analyses, have generally also found estimates of Fst to be much less than Wright's critical value [25%]. Genetic substructure does exist in humans, but there are no natural divisions in our species equivalent to biological races.
- The Princeton Guide to Evolution, 2017.
Quantitatively, one commonly used threshold is that two populations with sharp boundaries are considered to be different races if 25 percent or more of the genetic variability that they collectively share is found as between population differences.
- The Status of the Race Concept in Physical Anthropology
- Maglo, Koffi N., Tesfaye B. Mersha, and Lisa J. Martin. "Population Genomics and the Statistical Values of Race: An Interdisciplinary Perspective on the Biological Classification of Human Populations and Implications for Clinical Genetic Epidemiological Research." Frontiers in genetics 7 (2016).
- Genetic Similarities Within and Between Human Populations by D. J. Witherspoon et al. 2007. Genetics. 2007 May; 176(1): 351–359. doi:10.1534/genetics.106.067355.
- Johann Frederick Blumenbach. De generis humani varietate nativa, pp. XII-XLII. Gottingae : Vandenhoek et Ruprecht.
- Handley, L. Manica, A., Goudet, J., Balloux, F. (2007). "Going the Distance: Human Population Genetics in a Clinal World". Trends in Genetics. 23(9): 432-439.
- Clines, Clusters, and the Effect of Study Design on the Inference of Human Population Structure by Noah A Rosenberg et al. (2005) PLoS Genet. 1(6):e70. doi:10.1371/journal.pgen.0010070.
- The Genetic Structure and History of Africans and African Americans by Sarah A. Tishkoff (2009) Science 324(5930):1035–1044. doi:10.1126/science.1172257.
- Evidence for Gradients of Human Genetic Diversity Within and Among Continents by David Serra & Svante Pääbo (2004) Genome Research 14:1679-1685. doi:10.1101/gr.2529604.
- The Kenyan success genetic controversy by Ross Tucker (13 April 2013) The Science of Sport.
- [Pourquoi les Blancs courent moins vite by Jean-Philippe Leclaire (2012). Grasset. ISBN 2246785316.
- French book examines why black sprinters dominate the Olympics (2012-08-06) France 24.
- Craniometric similarities within and between human populations in comparison with neutral genetic data by A. Strauss and M. Hubbe. Hum. Biol. 2010 Jun;82(3):315-30. doi: 10.3378/027.082.0305.
- White Supremacist Finds Out His Ancestors Were Black by The Young Turks (Nov 13, 2013) The Trisha Goddard Show (archived copy).
- Barbujani, Guido, et al. An apportionment of human DNA diversity. Proceedings of the National Academy of Sciences 94.9 (1997): 4516-4519:
Zones of discontinuity in human gene frequency distributions are present, but the local gradients are so small that they can be identified only by simultaneously studying many loci using complex statistical techniques. In addition, such regions of relatively sharp genetic change do not surround large clusters of populations, on a continental or nearly continental scale. On the contrary, they occur irregularly, within continents and even within single countries.
See also Apportionment of Racial Diversity: a Review by Brown, R. A., Armelagos, G. J. (2001). Evolutionary Anthropology. 10(1):34-40. - Human Races: Classifying People vs. Understanding Diversity by Barbujani, G. (2005). Current Genomics. 6(4): 215-226:
"Classical population-genetics theory and empirical data show that large genetic differences among groups develop if reproductive barriers separate these groups. In the presence of reproductive barriers, that is, under isolation, genetic drift, affecting independently each group, will reduce the group’s internal variation (because the alleles that are lost are not reintroduced by gene flow) and will lead groups to diverge from each other (because different alleles are lost in each group and different mutations occur in each group); divergence will generate genetic discontinuities... between groups."
- Partitioning of Genetic Variation in Human Populations and the Concept of Race by Madrigal, L. & Barbujani, G. (2007). In: Crawford, M. (ed.). Anthropological Genetics. Cambridge University Press.
- Race, Ancestry, and Medicine by Feldman, M. W. & Lewontin, R. (2008). In: Koenig, B. A. (ed.). Revisiting Race in a Genomic Age. Rutgers University Press.
- Genetic Basis of Human Biodiversity: An Update by Guido Barbujani & Vincenza Colonna, in Biodiversity Hotspots, 2011. Springer, ISBN 9783642209918.
- The Apportionment of Human Diversity by Lewontin, R. (1972). Evolutionary Biology. 6: 381–398.
- Genetic differences within and between populations of the major human subgroups. by B. D. H. Latter (1980). Am. Natu. 116:220-237.
- Differences in the relative distribution of human gene diversity between electrophoretic and red and white cell antigen loci. by N. Ryman et al. Hum. Hered. 33:93-102.
- Relethford, John H. "Craniometric variation among modern human populations." American Journal of Physical Anthropology 95.1 (1994): 53-62.
- Relethford, J. H. (2002). "Apportionment of Global Human Genetic Diversity Based on Craniometrics and Skin Color". American Journal of Physical Anthropology. 118(4): 393-398.
- Microsatellite diversity and demographic history of modern humans. by L. B. Jorde et al. Proc. Natl. Acad. Sci. U.S.A. 94:3100-31003.
- Patterns of Human Diversity, within and among Continents, Inferred from Biallelic DNA Polymorphisms by Chiara Romualdi et al. Genome Res. 2002 Apr; 12(4): 602–612. doi:10.1101/gr.214902.
- Genetic structure of human populations by N. A. Rosenberg et al. Science. 2002 Dec 20;298(5602):2381-5.
- Genetic Variation Among World Populations: Inferences From 100 Alu Insertion Polymorphisms by W. Scott Watkins et al. Genome Res. 2003 Jul; 13(7): 1607–1618.
- Robustness of the inference of human population structure: A comparison of X-chromosomal and autosomal microsatellites by Sohini Ramachandran et al. Hum. Genomics. 2004; 1(2): 87–97. doi:10.1186/1479-7364-1-2-87.
- The genetic structure of human populations studied through short insertion-deletion polymorphisms by L. Bastos-Rodrigues et al. Ann. Hum. Genet. 2006 Sep;70(Pt 5):658-65.
- Worldwide human relationships inferred from genome-wide patterns of variation by J. Z. Li et al. Science. 2008 Feb 22;319(5866):1100-4. doi: 10.1126/science.1153717.
- A Population-Genetic Perspective on the Similarities and Differences among Worldwide Human Populations by Noah A. Rosenberg. Hum. Biol. 2011 Dec; 83(6): 659–684. doi:10.3378/027.083.0601.
- Global Geometric Morphometric Analyses of the Human Pelvis Reveal Substantial Neutral Population History Effects, Even across Sexes by Lia Betti et al. PLoS One. 2013; 8(2):e55909. doi:10.1371/journal.pone.0055909.
- On the New Biology of Race by Glasgow, J. M. (2003). The Journal of Philosophy. 100(9): 456-474.
- Unnaturalised racial naturalism by Adam Hochman, Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences, Volume 46, June 2014, Pages 79–87.
- Against the New Racial Naturalism by Adam Hochman (2013). The Journal of Philosophy. 110(6): 331-351:
The standard criterion for subspecies division is an FST estimate of or over 0.25.
andHowever, the new racial naturalism is not convincing. The continental clusters appealed to by race naturalists are arbitrary and superficial groupings, which should not be elevated to subspecies status. Moreover, the criteria applied to humans are not consistent with those used to define subspecies in nonhuman animals, and no rationale has been given for this differential treatment.
- Human Races: A Genetic and Evolutionary Perspective by Alan R. Templeton (1998). American Anthropologist 100(3):632-650.
- Geographic and Microgeographic Races by Marshall T. Newman. Current Anthropology, Vol. 4, No. 2 (Apr., 1963), pp. 189-207. (see Jean Hiernaux comments on pages 197-198).
- Stanford Encyclopedia of Philosophy (Race)
- Fearful Memories Passed Down to Mouse Descendants: Genetic imprint from traumatic experiences carries through at least two generations By Ewen Callaway, Scientific American (from Nature magazine on December 1, 2013).
- Epigenetics and the embodiment of race: developmental origins of US racial disparities in cardiovascular health by C. W. Kuzawa and E. Sweet. Am. J. Hum. Biol. 2009 Jan-Feb;21(1):2-15. doi:10.1002/ajhb.20822.
- Socioeconomic status modifies heritability of IQ in young children by Eric Turkheimer et al. 2003. Psychology Today vol. 14, no. 6, pp. 623-628.
- How Caucasoids got such big crania and why they shrank: from Morton to Rushton by Lieberman, L. (2001). Current Anthropology. 42(1): 69-95.
- Ethnicity and Race Overview by Dennis O'Neil, Palomar College.
- from PBS: Race — the power of an illusion — background readings Interview with Joseph Graves, Jr. (2003) PBS.
- Convergent adaptation of human lactase persistence in Africa and Europe by Sarah A Tishkoff et al. Nat. Genet. Jan 2007; 39(1): 31–40.
- Meta-analysis of associations between human brain volume and intelligence differences: How strong are they and what do they mean?
- Fact or Fiction: When It Comes to Intelligence, Does Brain Size Matter? What does brain size say about a creature's mental abilities? by Kayt Sukel (April 14, 2009) Scientific American.
- The Flynn Effect by Charles Graham (2001), revised by Jonathan Plucker (2002). See also None of the Above: What I.Q. doesn’t tell you about race by Malcolm Gladwell (December 17, 2007) The New Yorker; and The Flynn Effect and IQ Disparities Among Races, Ethnicities, and Nations: Are There Common Links? by Scott Barry Kaufman (August 23, 2010) Psychology Today.
- James R. Flynn. Searching for Justice: The Discovery of IQ Gains Over Time by James R. Flynn. American Psychologist, vol. 54, no. 1, Jan. 1999, pp. 5-20.
- Researchers Say Human Brain Is Still Evolving by Nicholas Wade (September 8, 2005) The New York Times.
- The genetics of lactase persistence by Alison Campbell (June 25, 2010 2:19 PM). The University of Waikato.
- Is it wrong to note 100m winners are always black?, BBC, 2011
- A True Champion Vs. The 'Great White Hope', commentary by John Ridley (July 2, 201012:00 AM ET) NPR.
- Why don't black Americans swim? by Finlo Rohrer (3 September 2010) BBC.
- Kenyan siblings in pursuit of 2012 gold by Ben Oakley (Page last updated at 14:49 GMT, Friday, 6 August 2010 15:49 UK) BBC.
- Why Chinese Men Stink At Soccer by Gordon G. Chang (7/07/2010 @ 12:38PM ) Forbes.
- Why Aren't Asian-Americans Getting Their 'One Shining Moment'? by Gene Demby (April 2, 20147:00 AM ET) NPR.
- Tennis: Why So Few Blacks? by Bud Withers (July 8, 1973) Eugene Register-Guard.
- White Man Can't Jump: The Science Behind Stereotypes by Nikki Tucker (Sep 5, 2012 12:12 PM) Medical Daily.
- See the Wikipedia article on All-time Olympic Games medal table.
- Scientists try to answer why Dutch people are so tall (Tuesday 7 April 2015 21.21 EDT) The Guardian (via Agence France-Presse).
- See the Wikipedia article on Race and sports.
- Race, genes, and sports by William Saletan (Dec. 5 2008 7:30 AM) Slate.
- Race-Based Medicine: A Recipe for Controversy: Is race-based medicine a boon or boondoggle?, Editorial (Jul 31, 2007) Scientific American
- Genetic Anthropology, Ancestry, and Ancient Human Migration (Human Genome Project Information Archive)
- What We Do and Don't Know About 'Race', 'Ethnicity', Genetics, and Health at the Dawn of the Genome Era. by Francis S. Collins. Nature Genetics Supplement, vol. 36, no. 11, Nov. 2004.
- "Genetic Structure, Self-Identified Race/Ethnicity, and Confounding in Case-Control Association Studies" by Hua Tang, et al. Am. J. Hum. Genet. 2005 Feb; 76(2): 268–275.
- "Defining Human Differences in Biomedicine" by Maggie Brown PLoS Medicine. 2007. 4(9): e228. doi:10.1371/journal.pmed.0040288.
- S.D.J. Pena. "The fallacy of racial pharmacogenomics". Brazilian Journal of Medical and Biological Research (2011) 44: 268-275
- Race and Gene Studies: What Difference Makes a Difference? by Larry Adelman (2003) California Newsreel.
- Implications of biogeography of human populations for 'race' and medicine by Sarah A. Tishkoff & Kenneth K. Kidd, Nature Genetics 36, S21-S27 (2004). doi:10.1038/ng1438.
- Do Races Differ? Not Really, DNA Shows Natalie (August 22, 2000) New York Times.
- (Templeton, 2019, p. 443
- Confusions About Human Races by R. C. Lewontin (Jun 07, 2006). Is Race Real? a web forum organization hoste by the Social Science Research Council.
- Prag, J., Neave, R. (1997). Making Faces: Using Forensic and Archaeological Evidence. Trustees of the British Museum. p. 34.
- The Human Skeleton in Forensic Medicine by Wilton Marion Krogman & Mehmet Yaşar İşcan (1986) Charles C. Thomas. 2nd edition.
- Forensic Anthropology and the Concept of Race: If Races Don't Exist, Why Are Forensic Anthropologists So Good at Identifying Them? by Sauer, N. J. (1992). Social Science & Medicine. 34(2):107-111. doi:10.1016/0277-9536(92)90086-6.
- Cranial Nonmetric Variation and Estimating Ancestry by Joseph T. Hefner (2009) J. Forensic Sci. 54(5):985-995. doi:10.1111/j.1556-4029.2009.01118.x.
- The Human Skeleton in Forensic Medicine by Mehmet Yaşar İşcan & Maryna Steyn (2013) Charles C. Thomas. 3rd edition. ISBN 0398088780.
- Smay, D., Armelagos, G. (2000). "Galileo Wept: a Critical Assessment of the Use of Race in Forensic Anthropology (pdf), (dotx)". Transforming Anthropology. 9(2): 19-29.
- Heard, A. N. (2008). "Non-Metric Assessment of Southeast and Northeast Asian Ancestry in the Forensic Context". M.Sc. Thesis. Michigan State University.
Recently, physical anthropologists have noted that traits considered to be characteristic of the classical Mongoloid group were not derived from studies encompassing all of the populations that would be classified as Mongoloid.
- An evaluation of race and sex identification from cranial measurements by Walter H. Birkby. Am. J. Phys. Anthropol. 24:21–27, 1966.
- Is It Possible to Escape Racial Typology in Forensic Identification? by Albanese, J., Saunders, S. R. (2006). In: Schmitt, A., Pinheiro, J. (eds.). Forensic Anthropology and Medicine. Humana.
- Can Ancestry be Consistently Determined From the Skeleton? by Sierp, I., Henneberg, M. (2015). Anthropological Review. 78(1): 21-31.
- "China". In: Spencer, F. (ed.). History of Physical Anthropology (vol. 1). New York: Garland.
- Ling, J. Y. K., Wong, R. W. K. (2008). "Incisal Morphology of Southern Chinese". The Open Anthropology Journal. 1: 19-25.
- Region Does Not Mean Race: Reality Versus Convention in Forensic Anthropology by Brace, C. L. (1995). Journal of Forensic Sciences. 40(2): 171-175.
- "Ten Facts about Human Variation" by Jonathan Marks, in Human Evolutionary Biology, edited by Michael P. Muehlenbein (2010). Cambridge University Press. ISBN 052170510X.
- Evidence for Gradients of Human Genetic Diversity Within and Among Continents by David David Serre & Svante Pääbo. Genome Research 2004. 14: 1679-1685. doi:10.1101/gr.2529604.
- Relethford, John. "Global patterns of isolation by distance based on genetic and morphological data." Human Biology 76.4 (2004): 499-513.
- Clines Without Classes: How to Make Sense of Human Variation by Fujimura, J. H., Bolnick, D. A., Rajagopalan, R., Kaufman, J. S., Lewontin, R. C. (2014). Sociological Theory. 32(3): 208-227.
- Statement on Race, UNESCO:
Specifically, between human populations, including smaller groupings, genetic differences may be detected. These differences tend to increase with geographic distance, but the basic genetic variation between populations is much less prominent. This means that human genetic diversity is only gradual and presents no major discontinuity between populations.
- Clines, Clusters, and the Effect of Study Design on the Inference of Human Population Structure by Noah A. Rosenberg, et al. PLOS Genetics (December 9, 2005) doi:10.1371/journal.pgen.0010070.
- "Zones of sudden genetic variation exist, often in correspondence with barriers to gene flow, both physical and cultural, but they almost never completely surround a region. Over much of the planet, clinal variation is the rule, with genetic characteristics of the various regions changing gradually into one another." ("Genetic Variances Between Human Groups" by Guido Barbujani & Giulietta Di Benedetto, in Genes, Fossils, and Behaviour, edited by Peter Donnelly & Robert A. Foley (2001). IOS Press. ISBN 9051994494.)
- "The genetic exchanges occurred in the course of the frequent contacts have resulted in a smooth, continuous variation of many genetic parameters. As a consequence, zones of sharp genetic change are not the rule, but the exception; most human populations are not surrounded by clear genetic boundaries." (Genetic Basis of Human Biodiversity: An Update by Guido Barbujani & Vincenza Colonna, in Biodiversity Hotspots, 2011. Springer, ISBN 9783642209918.)
- Marks, 2010.
- Richard Lewontin, The Genetic Basis of Evolutionary Change (1974)
- The Rational View on Race? by Sean Last (May 18, 2015) (comment by 8theist, ca. 2015) The Right Stuff.
- Smedley, Audrey, and Brian D. Smedley. "Race as biology is fiction, racism as a social problem is real: Anthropological and historical perspectives on the social construction of race." American Psychologist 60.1 (2005): 16.
- In Our Own Voices: African American Stories of Oppression, Survival and Recovery in the Mental Health System by Vanessa Jackson, pp. 1-36, 4-8.
- Eggs, eggplants and eggheads: a rejoinder to Rushton by Weizmann, F., Wiener, et al. (1991). Canadian Psychology, 32 (1), 43-50 doi:10.1037/h0078958.
- Rushton’s r–K life history theory of race differences in penis length and circumference examined in 113 populations by Richard Lynn. Personality and Individual Differences. Volume 55, Issue 3, July 2013, Pages 261–266. doi:10.1016/j.paid.2012.02.016.
- The Penile Economics of Ethnicity (March 3, 2012) Ethnic Muse.
- World Penis Average Size Studies Database
- WorldPenisSize: analysis of listed sources (May 1, 2012) Ethnic Muse.
- Racism Resurgent: How Media Let The Bell Curve's Pseudo-Science Define the Agenda on Race by Jim Naureckas, Fairness in Accuracy and Reporting, January/February 1995.
- Race Differences in Intelligence: A Global Perspective by Richard Lynn (1991). Mankind Quarterly, 31(3):255-296.
- AmRen Conference Held in Tennessee by Henry Wolff (March 19, 2012) American Renaissance.
- Is Size important? by Donald I. Templer (2002). CeShore, ISBN 1585010324.