Anchiornithidae
Anchiornithidae ("near birds") is a family of eumaniraptorans which could be the basalmost family of birds (in the general sense) in the clade Avialae.[1] Anchiornithids have been classified at varying positions in the maniraptoran tree, with some scientists classifying them as a distinct family, a basal subfamily of Troodontidae,[2][3][4][5] members of Archaeopterygidae,[2][6] or an assemblage of dinosaurs that are an evolutionary grade within Avialae[7] or Paraves.[8]
| Anchiornithidae | |
|---|---|
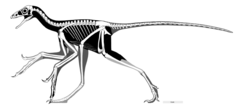 | |
| Skeletal restoration of Anchiornis huxleyi by Scott Hartman, 2017 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Avialae (?) |
| Family: | †Anchiornithidae Xu et al. 2016 sensu Foth & Rauhut, 2017 |
| Type species | |
| †Anchiornis huxleyi Xu et al., 2009 | |
| Genera | |
| Synonyms | |
| |
Description
Anchiornithids share many general features with other Paraves and early avialans. They were small and lightly-built feathered carnivores, similar in biology to Archaeopteryx, early dromaeosaurids like Microraptor, and particularly troodontids. They are almost exclusively known from Late Jurassic Chinese deposits, although Ostromia was discovered in Germany and Yixianosaurus (a putative member of the group only known from forelimbs) is believed to hail from the early Cretaceous. Most had long legs, arms, and hands, although some (Eosinopteryx) had slightly reduced forelimbs.[9]
Feathering

Although it is practically certain that every anchiornithid possessed advanced pennaceous feathers, there is still much variety in feathering between genera (or between individuals in the case of numerous genera such as Anchiornis). Most had vaned tail feathers forming a frond-shaped tail, with the tails feathers of Caihong being particularly long and in some cases asymmetrical.[10] However, Eosinopteryx, Serikornis, and Aurornis were preserved with short and downy tail feathering. Some studies on the body feathering of anchiornithids indicate that the feathers were pennaceous, but seemingly lacked barbules, making them "shaggy" or "silky" in life.[11][8]
Long pennaceous feathers were present on the arms of most anchiornithids. However, these feathers were slender, symmetrical, and unspecialized, probably useless for flight. They formed rows which were attached directly to a large fleshy propatagium connecting the upper and lower arm.[12][8]
Most anchiornithids also had dense feathering extending down their legs. A few had short leg feathering, but most (Anchiornis and Pedopenna, for example) had very long pennaceous feathers on their legs, giving them the moniker of "four-winged dinosaurs", a trait also shared by microraptorians. Eosinopteryx seemingly lacked any sort of feathers on the lower part of its legs, but its close relative Serikornis possessed both plumaceous (downy) feathers extending onto its toes as well as pennaceous feathers further up the leg.[9][8]
Distinguishing features
Foth and Rauhut (2017) established several diagnostic features present in anchiornithids:[1]
- Nutrient foramina on the dentary are placed in a deep groove (also in most troodontids and some other coelurosaurs);
- Anterior dentary teeth which are smaller, more numerous, and more closely spaced than those in the middle of the tooth row (also in most troodontids);
- The front edge of the acromion margin of the scapula is bent or hooked outwards (also in several oviraptorosaurs and more derived avialans);
- The inside surface of proximal part of the fibula is flat (also in alvarezsaurids, therizinosauroids and derived avialans);
- Fan-shaped posterior dorsal neural spines (also in compsognathids and some derived avialans);
- Extensive large pennaceous feathers on the foot and ankle (also in Microraptor and Sapeornis).
Systematic history
In 2015 Chatterjee created Tetrapterygidae in the second edition of his book The Rise of Birds: 225 Million Years of Evolution, where he included Xiaotingia, Aurornis, Anchiornis, and even Microraptor; together they were proposed to be the sister group of the Avialae.[13] However this family is invalid as must include the genus Tetrapteryx, which is the junior synonym of Grus – therefore Tetrapterygidae is a junior synonym of Gruidae.[14]
In their description of Wiehenvenator Rauhut and colleagues had informally called the group as "Anchiornithosaurs" which they placed outside of Avialae.[15]
The clade was originally named as "Anchiornithinae" by Xu et al. (2016) and defined as for "the most inclusive clade including Anchiornis but not Archaeopteryx, Gallus, Troodon, Dromaeosaurus, Unenlagia, or Epidexipteryx".[16]
In 2017 Foth and Rauhut in their re-evaluation of the Haarlem Archaeopteryx specimen (which they classified it in its own distinct genus Ostromia[1]) found that the anchiornithids are a distinct family closer to the ancestry of birds.[1] They provided their own definition of Anchiornithidae as "as all maniraptoran theropods that are more closely related to Anchiornis huxleyi than to Passer domesticus, Archaeopteryx lithographica, Dromaeosaurus albertensis, Troodon formosus, or Oviraptor philoceratops."[1]
During the description of Halszkaraptor, Cau et al. (2017) incorporated many putative anchiornithids into two different large-scale phylogenetic analyses. The first analysis was a comprehensive study of theropod dinosaurs originally designed by Lee et al. for a 2014 paper on miniaturization in theropods leading up to the evolution of birds. Cau et al.'s usage of this first analysis found support for Anchiornithidae being a distinctive family of avialans. The strict consensus tree of the first analysis is given below:[17]
| Avialae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The second analysis was first used in a different paper on theropod size published by Brusatte et al. in 2014. This analysis (which was updated by Cau et al. during a 2015 study on the affinities of Balaur bondoc[18]) focused specifically on coelurosaurs and found that anchiornithids (represented only by Anchiornis, Xiaotingia, Aurornis, and Eosinopteryx in the analysis) were troodontids rather than avialans, in contrast to the first analysis.[17]
The description of Caihong by Hu et al. (2018) also implemented the Brusatte analysis and found the same result. However, this study also implemented an analysis performed by Xu et al. (2015) during the description of Yi qi. This analysis placed anchiornithids (or as the study calls them, members of "Anchiorninae") either as troodontids or unresolved paravians, depending on whether parsimony or bootstrap analyses are used.[19]
An analysis used in the description of the Jurassic bird Alcmonavis by Rauhut et al. (2019) recovered anchiornithids (represented in the analysis by Eosinopteryx, Anchiornis, and Ostromia) as the most basal avialans. Xiaotingia and Pedopenna were placed as more advanced avialans closer to Archaeopteryx.[20]
In 2019 with the description of the Late Jurassic genus Hesperornithoides, Hartman et al., using every named Mesozoic maniraptoromorph (with the addition of 28 unnamed specimens), which they scored 700 characters and 501 operational taxonomic units, found that most of the anchiornithids are members of Archaeopterygidae, with only Xiaotingia and Yixianosaurus being classified as a troodontid and a dromaeosaurid respectfully, Pedopenna found in many possible positions within the Paraves phylogeny, and Ostromia described too late before adding into the analysis.[6] Below is their phylogeny:
| Archaeopterygidae |
| ||||||||||||||||||||||||||||||
The cladogram below shows the results of the phylogenetic analysis by Cau (2020).[21]
| |||||||||||||||||||||||||||||||||||||||||||||||||
Palaeoecology
A good majority of the known anchiornithid fossils are have been recovered from the Tiaojishan Formation in Liaoning, China dating back to 160 million years.[22][23] The climate during this period of time would have been subtropical to temperate, warm and humid based on the plant life present in the Tiaojishan Formation.[24] This environment was dominated plant by gymnosperm trees. There were ginkgopsids like Ginkoites, Ginkgo, Baiera, Czekanowskia, and Phoenicopsis. There were also conifers like Pityophyllum, Rhipidiocladus, Elatocladus, Schizolepis, and Podozamites. Also, Lycopsids like Lycopodites and Sellaginellities, horsetails (Sphenopsida) like Equisetum, cycads like Anomozamites, and ferns (Filicopsida) like Todites and Coniopteris.[25] The only Chinese anchiornithid to not have been discovered in the Tiaojishan Formation is Yixianosaurus longimanus, which was found in the 125 million-year-old Early Cretaceous Yixian Formation, in the same area. Only one genus of anchiornithid has been found outside of China: Ostromia, which is found in the Painten Formation from Riedenburg, Bavaria, Germany.[1]
References
- Foth, C.; Rauhut, O. W. M. (2017). "Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs". BMC Evolutionary Biology. 17 (1): 236. doi:10.1186/s12862-017-1076-y. PMC 5712154. PMID 29197327.
- Xing Xu, Hailu You, Kai Du and Fenglu Han (28 July 2011). "An Archaeopteryx-like theropod from China and the origin of Avialae" (PDF). Nature. 475 (7357): 465–470. doi:10.1038/nature10288. PMID 21796204. Archived from the original (PDF) on 20 December 2016. Retrieved 6 December 2017.CS1 maint: uses authors parameter (link)
- Lee, M. S. Y. and Worthy, T. H. (2011). "Likelihood reinstates Archaeopteryx as a primitive bird". Biology Letters. 8 (2): 299–303. doi:10.1098/rsbl.2011.0884. PMC 3297401. PMID 22031726.CS1 maint: multiple names: authors list (link)
- Stephen L. Brusatte; Graeme T. Lloyd; Steve C. Wang; Mark A. Norell (2014). "Gradual assembly of avian body plan culminated in rapid rates of evolution across the dinosaur-bird transition". Current Biology. 24 (20): 2386–2392. doi:10.1016/j.cub.2014.08.034. PMID 25264248.
- Cai-zhi Shen; Bo Zhao; Chun-ling Gao; Jun-chang Lü; Martin Kundrát (2017). "A New Troodontid Dinosaur (Liaoningvenator curriei gen. et sp. nov.) from the Early Cretaceous Yixian Formation in Western Liaoning Province". Acta Geoscientica Sinica. 38 (3): 359–371. doi:10.3975/cagsb.2017.03.06.
- Hartman, Scott; Mortimer, Mickey; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. (2019). "A new paravian dinosaur from the Late Jurassic of North America supports a late acquisition of avian flight". PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906.
- Wang, M.; Wang, X.; Wang, Y.; Zhou, Z. (2016). "A new basal bird from China with implications for morphological diversity in early birds". Scientific Reports. 6: 19700. doi:10.1038/srep19700. PMC 4726217. PMID 26806355.
- Lefèvre, Ulysse; Cau, Andrea; Cincotta, Aude; Hu, Dongyu; Chinsamy, Anusuya; Escuillié, François; Godefroit, Pascal (2017). "A new Jurassic theropod from China documents a transitional step in the macrostructure of feathers". The Science of Nature. 104 (9–10): 74. Bibcode:2017SciNa.104...74L. doi:10.1007/s00114-017-1496-y. PMID 28831510.
- Godefroit, Pascal; Demuynck, Helena; Dyke, Gareth; Hu, Dongyu; Escuillié, François; Claeys, Philippe (22 January 2013). "Reduced plumage and flight ability of a new Jurassic paravian theropod from China". Nature Communications. 4: 1394. doi:10.1038/ncomms2389. PMID 23340434.
- Dongyu Hu; Julia A. Clarke; Chad M. Eliason; Rui Qiu; Quanguo Li; Matthew D. Shawkey; Cuilin Zhao; Liliana D’Alba; Jinkai Jiang; Xing Xu (2018). "A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution". Nature Communications. 9 (1): Article number 217. Bibcode:2018NatCo...9..217H. doi:10.1038/s41467-017-02515-y. PMC 5768872. PMID 29335537.
- Saitta, Evan T.; Gelernter, Rebecca; Vinther, Jakob (2018). "Additional information on the primitive contour and wing feathering of paravian dinosaurs". Palaeontology. 61 (2): 273–288. doi:10.1111/pala.12342. hdl:1983/61351c6d-1517-4101-bac8-50cbb733761d. ISSN 1475-4983.
- Wang, Xiaoli; Pittman, Michael; Zheng, Xiaoting; Kaye, Thomas G.; Falk, Amanda R.; Hartman, Scott A.; Xu, Xing (1 March 2017). "Basal paravian functional anatomy illuminated by high-detail body outline". Nature Communications. 8: 14576. Bibcode:2017NatCo...814576W. doi:10.1038/ncomms14576. PMC 5339877. PMID 28248287.
- Chatterjee, S. (2015). The rise of birds: 225 million years of evolution. Johns Hopkins University Press, 45–48.
- Matthew Martyniuk (24 May 2015). "The Crane and the Microraptor". DinoGoss: A blog about stem-birds. Archived from the original on 16 June 2015.
- Rauhut, O.W.M., Hübner T.R., and Lanser, K., 2015, "A new theropod dinosaur from the late Middle Jurassic of Germany and theropod faunal turnover during the Jurassic", Libro de resúmenes del V Congreso Latinoamericano de Paleontología de Vertebrados. 62
- Xu; et al. (2016). "An Updated Review of the Middle-Late Jurassic Yanliao Biota: Chronology, Taphonomy, Paleontology and Paleoecology". Acta Geologica Sinica. 90 (6): 2229–2243. doi:10.1111/1755-6724.13033.
- Cau, A.; Beyrand, V.; Voeten, D.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P.; Godefroit, P. (2017). "Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs". Nature. 552 (7685): 395–399. Bibcode:2017Natur.552..395C. doi:10.1038/nature24679. PMID 29211712.
- Cau, Andrea; Brougham, Tom; Naish, Darren (18 June 2015). "The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropodBalaur bondoc(Dinosauria, Maniraptora): dromaeosaurid or flightless bird?". PeerJ. 3: e1032. doi:10.7717/peerj.1032. ISSN 2167-8359. PMC 4476167. PMID 26157616.
- Xu, Xing; Jiang, Jinkai; D’Alba, Liliana; Zhao, Cuilin; Matthew D. Shawkey; Li, Quanguo; Qiu, Rui; Eliason, Chad M.; Clarke, Julia A. (15 January 2018). "A bony-crested Jurassic dinosaur with evidence of iridescent plumage highlights complexity in early paravian evolution". Nature Communications. 9 (1): 217. Bibcode:2018NatCo...9..217H. doi:10.1038/s41467-017-02515-y. ISSN 2041-1723. PMC 5768872. PMID 29335537.
- Rauhut, O.; Tischlinger, H.; Foth, C. (14 May 2019). "A non-archaeopterygid avialan theropod from the Late Jurassic of southern Germany". eLife. 8: e43789. doi:10.7554/eLife.43789. ISSN 2050-084X. PMC 6516837. PMID 31084702.
- Cau, Andrea (25 February 2020). "The body plan of Halszkaraptor escuilliei (Dinosauria, Theropoda) is not a transitional form along the evolution of dromaeosaurid hypercarnivory". PeerJ. 8: e8672. doi:10.7717/peerj.8672. ISSN 2167-8359. PMC 7047864. PMID 32140312.
- Zhou, Z.-H.; Wang, Y. (2017). "Vertebrate assemblages of the Jurassic Yanliao Biota and the Early Cretaceous Jehol Biota: Comparisons and implications". Paleoworld. 26 (2): 241. doi:10.1016/j.palwor.2017.01.002.
- Sullivan, C.; Wang, Y.; Hone, D.W.E.; Wang, Y.; Xu, X.; Zhang, F. (2014). "The vertebrates of the Jurassic Daohugou Biota of northeastern China". Journal of Vertebrate Paleontology. 34 (2): 243–280. doi:10.1080/02724634.2013.787316.
- Wang, Y.; Ken, S.; Zhang, W.; Zheng, S. (2006). "Biodiversity and palaeoclimate of the Middle Jurassic floras from the Tiaojishan Formation in western Liaoning, China". Progress in Natural Science. 16 (1): 222–230. doi:10.1080/10020070612330087.
- Zhang, Kuiyan; Yang, Ding; Ren, Dong (2006). "The first snipe fly (Diptera: Rhagionidae) from the Middle Jurassic of Inner Mongolia, China". Zootaxa. 1134: 51–57. doi:10.11646/zootaxa.1134.1.3.



.png)





.png)


