PSMD2
26S proteasome non-ATPase regulatory subunit 2, also as known as 26S Proteasome Regulatory Subunit Rpn1 (systematic nomenclature), is an enzyme that in humans is encoded by the PSMD2 gene.[5][6]

Structure
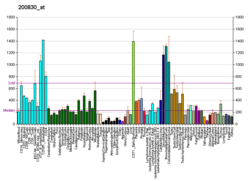
Gene expression
The gene PSMD2 encodes a non-ATPase subunit of the 19S regulator base, which is responsible for substrate recognition and binding.[6] The gene PSMD2 encodes one of the non-ATPase subunits of the 19S regulator lid. In addition to participation in proteasome function, this subunit may also participate in the TNF signalling pathway since it interacts with the tumor necrosis factor type 1 receptor. A pseudogene has been identified on chromosome 1.[6] The human PSMD2 gene has 23 exons and locates at chromosome band 3q27.1. The human protein 26S proteasome non-ATPase regulatory subunit 2 is 100 kDa in size and composed of 909 amino acids. The calculated theoretical pI of this protein is 5.10. Two expression isoforms are generated by alternative splicing, in which either 1-130 or 1-163 of the amino acid sequence is missing.
Complex assembly
26S proteasome complex is usually consisted of a 20S core particle (CP, or 20S proteasome) and one or two 19S regulatory particles (RP, or 19S proteasome) on either one side or both side of the barrel-shaped 20S. The CP and RPs pertain distinct structural characteristics and biological functions. In brief, 20S sub complex presents three types proteolytic activities, including caspase-like, trypsin-like, and chymotrypsin-like activities. These proteolytic active sites located in the inner side of a chamber formed by 4 stacked rings of 20S subunits, preventing random protein-enzyme encounter and uncontrolled protein degradation. The 19S regulatory particles can recognize ubiquitin-labeled protein as degradation substrate, unfold the protein to linear, open the gate of 20S core particle, and guide the substate into the proteolytic chamber. To meet such functional complexity, 19S regulatory particle contains at least 18 constitutive subunits. These subunits can be categorized into two classes based on the ATP dependence of subunits, ATP-dependent subunits and ATP-independent subunits. According to the protein interaction and topological characteristics of this multisubunit complex, the 19S regulatory particle is composed of a base and a lid subcomplex. The base consists of a ring of six AAA ATPases (Subunit Rpt1-6, systematic nomenclature) and four non-ATPase subunits (Rpn1, Rpn2, Rpn10, and Rpn13). Thus, Protein 26S proteasome non-ATPase regulatory subunit 2 (Rpn1) is an essential component of forming the base subcomplex of 19S regulatory particle. Traditionally, Rpn1 and Rpn2 were considered residing at the center of base sub complex and surrounded by six AAA ATPases (Rpt 1-6). However, recent investigation provides an alternative structure of 19S base via an integrative approach combining data from cryoelectron microscopy, X-ray crystallography, residue-specific chemical cross-linking, and several proteomics techniques. Rpn2 is rigid protein located on the side of ATPase ring, supporting as the connection between the lid and base. Rpn1 is conformationally variable, positioned at the periphery of the ATPase ring. The ubiquitin receptors Rpn10 and Rpn13 are located further in the distal part of the 19S complex, indicating that they were recruited to the complex late during the assembly process.[7]
Function
As the degradation machinery that is responsible for ~70% of intracellular proteolysis,[8] proteasome complex (26S proteasome) plays a critical roles in maintaining the homeostasis of cellular proteome. Accordingly, misfolded proteins and damaged protein need to be continuously removed to recycle amino acids for new synthesis; in parallel, some key regulatory proteins fulfill their biological functions via selective degradation; furthermore, proteins are digested into peptides for MHC class I antigen presentation. To meet such complicated demands in biological process via spatial and temporal proteolysis, protein substrates have to be recognized, recruited, and eventually hydrolyzed in a well controlled fashion. Thus, 19S regulatory particle pertains a series of important capabilities to address these functional challenges. To recognize protein as designated substrate, 19S complex has subunits that are capable to recognize proteins with a special degradative tag, the ubiquitinylation. It also have subunits that can bind with nucleotides (e.g., ATPs) in order to facilitate the association between 19S and 20S particles, as well as to cause confirmation changes of alpha subunit C-terminals that form the substate entrance of 20S complex. Rpn1 is one essential subunit of 19S regulatory particle and it forms the core of the "base" subcomplex. It offers a docking position for another 19S subunit Rpn10 at its central solenoid portion, although such association with Rpn10 is stabilized by a third subunit, Rpn2.[9] Besides its critical roles in 19S complex assembly, Rpn2 also provides docking positions for shuttles of ubiqitinylated substrate trafficking. The majority of shuttles attach to the proteasome via a ubiquitin-like domain (UBL) while they unload the substrate cargo at a C-terminal polyubiquitin-binding domain(s). Recent investigation by Glickman et al. identified that two shuttle proteins, Rad23 and Dsk2, dock at two different receptor sites embedded within subunit Rpn1.[9]
Clinical significance
The proteasome and its subunits are of clinical significance for at least two reasons: (1) a compromised complex assembly or a dysfunctional proteasome can be associated with the underlying pathophysiology of specific diseases, and (2) they can be exploited as drug targets for therapeutic interventions. More recently, more effort has been made to consider the proteasome for the development of novel diagnostic markers and strategies. An improved and comprehensive understanding of the pathophysiology of the proteasome should lead to clinical applications in the future.
The proteasomes form a pivotal component for the ubiquitin–proteasome system (UPS) [10] and corresponding cellular Protein Quality Control (PQC). Protein ubiquitination and subsequent proteolysis and degradation by the proteasome are important mechanisms in the regulation of the cell cycle, cell growth and differentiation, gene transcription, signal transduction and apoptosis.[11] Subsequently, a compromised proteasome complex assembly and function lead to reduced proteolytic activities and the accumulation of damaged or misfolded protein species. Such protein accumulation may contribute to the pathogenesis and phenotypic characteristics in neurodegenerative diseases,[12][13] cardiovascular diseases,[14][15][16] inflammatory responses and autoimmune diseases,[17] and systemic DNA damage responses leading to malignancies.[18]
Several experimental and clinical studies have indicated that aberrations and deregulations of the UPS contribute to the pathogenesis of several neurodegenerative and myodegenerative disorders, including Alzheimer's disease,[19] Parkinson's disease[20] and Pick's disease,[21] Amyotrophic lateral sclerosis (ALS),[21] Huntington's disease,[20] Creutzfeldt–Jakob disease,[22] and motor neuron diseases, polyglutamine (PolyQ) diseases, Muscular dystrophies[23] and several rare forms of neurodegenerative diseases associated with dementia.[24] As part of the ubiquitin–proteasome system (UPS), the proteasome maintains cardiac protein homeostasis and thus plays a significant role in cardiac ischemic injury,[25] ventricular hypertrophy[26] and Heart failure.[27] Additionally, evidence is accumulating that the UPS plays an essential role in malignant transformation. UPS proteolysis plays a major role in responses of cancer cells to stimulatory signals that are critical for the development of cancer. Accordingly, gene expression by degradation of transcription factors, such as p53, c-jun, c-Fos, NF-κB, c-Myc, HIF-1α, MATα2, STAT3, sterol-regulated element-binding proteins and androgen receptors are all controlled by the UPS and thus involved in the development of various malignancies.[28] Moreover, the UPS regulates the degradation of tumor suppressor gene products such as adenomatous polyposis coli (APC) in colorectal cancer, retinoblastoma (Rb). and von Hippel–Lindau tumor suppressor (VHL), as well as a number of proto-oncogenes (Raf, Myc, Myb, Rel, Src, Mos, ABL). The UPS is also involved in the regulation of inflammatory responses. This activity is usually attributed to the role of proteasomes in the activation of NF-κB which further regulates the expression of pro inflammatory cytokines such as TNF-α, IL-β, IL-8, adhesion molecules (ICAM-1, VCAM-1, P-selectin) and prostaglandins and nitric oxide (NO).[17] Additionally, the UPS also plays a role in inflammatory responses as regulators of leukocyte proliferation, mainly through proteolysis of cyclines and the degradation of CDK inhibitors.[29] Lastly, autoimmune disease patients with SLE, Sjögren syndrome and rheumatoid arthritis (RA) predominantly exhibit circulating proteasomes which can be applied as clinical biomarkers.[30]
The protein 26S proteasome non-ATPase regulatory subunit 2 (Rpn1) which is encoded by PSMD2 has been identified as an important constituent of a signature associated with the acquisition of metastatic phenotype and poor prognosis in lung cancers.[31] It was found that knockdown of PSMD2 decreased proteasome activity, and induced growth inhibition and apoptosis in lung cancer cell lines. These effects of siRNA-mediated PSMD2 inhibition were associated with changes in the balance between phosphorylated AKT and p38, as well as with the induction of p21. In addition, patients with higher PSMD2 expression indicated a poorer prognosis and a small fraction of lung cancer specimens carried increased copies of PSMD2. Notably, findings illustrate that lung adenocarcinomas can be divided into two main groups; those with and without general upregulation of proteasome pathway genes including PSMD2.[31]
Interactions
PSMD2 has been shown to interact with TNFRSF1A[32][33] and PSMC1.[34][35]
References
- GRCh38: Ensembl release 89: ENSG00000175166 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000006998 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Tsurumi C, Shimizu Y, Saeki M, Kato S, Demartino GN, Slaughter CA, Fujimuro M, Yokosawa H, Yamasaki M, Hendil KB, Toh-e A, Tanahashi N, Tanaka K (October 1996). "cDNA cloning and functional analysis of the p97 subunit of the 26S proteasome, a polypeptide identical to the type-1 tumor-necrosis-factor-receptor-associated protein-2/55.11". Eur J Biochem. 239 (3): 912–21. doi:10.1111/j.1432-1033.1996.0912u.x. PMID 8774743.
- "Entrez Gene: PSMD2 proteasome (prosome, macropain) 26S subunit, non-ATPase, 2".
- Lasker K, Förster F, Bohn S, Walzthoeni T, Villa E, Unverdorben P, Beck F, Aebersold R, Sali A, Baumeister W (Jan 2012). "Molecular architecture of the 26S proteasome holocomplex determined by an integrative approach". Proceedings of the National Academy of Sciences of the United States of America. 109 (5): 1380–7. Bibcode:2012PNAS..109.1380L. doi:10.1073/pnas.1120559109. PMC 3277140. PMID 22307589.
- Rock KL, Gramm C, Rothstein L, Clark K, Stein R, Dick L, Hwang D, Goldberg AL (Sep 1994). "Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules". Cell. 78 (5): 761–71. doi:10.1016/s0092-8674(94)90462-6. PMID 8087844.
- Rosenzweig R, Bronner V, Zhang D, Fushman D, Glickman MH (Apr 2012). "Rpn1 and Rpn2 coordinate ubiquitin processing factors at proteasome". The Journal of Biological Chemistry. 287 (18): 14659–71. doi:10.1074/jbc.M111.316323. PMC 3340268. PMID 22318722.
- Kleiger G, Mayor T (Jun 2014). "Perilous journey: a tour of the ubiquitin–proteasome system". Trends in Cell Biology. 24 (6): 352–9. doi:10.1016/j.tcb.2013.12.003. PMC 4037451. PMID 24457024.
- Goldberg AL, Stein R, Adams J (Aug 1995). "New insights into proteasome function: from archaebacteria to drug development". Chemistry & Biology. 2 (8): 503–8. doi:10.1016/1074-5521(95)90182-5. PMID 9383453.
- Sulistio YA, Heese K (Jan 2015). "The Ubiquitin-Proteasome System and Molecular Chaperone Deregulation in Alzheimer's Disease". Molecular Neurobiology. 53 (2): 905–31. doi:10.1007/s12035-014-9063-4. PMID 25561438.
- Ortega Z, Lucas JJ (2014). "Ubiquitin-proteasome system involvement in Huntington's disease". Frontiers in Molecular Neuroscience. 7: 77. doi:10.3389/fnmol.2014.00077. PMC 4179678. PMID 25324717.
- Sandri M, Robbins J (Jun 2014). "Proteotoxicity: an underappreciated pathology in cardiac disease". Journal of Molecular and Cellular Cardiology. 71: 3–10. doi:10.1016/j.yjmcc.2013.12.015. PMC 4011959. PMID 24380730.
- Drews O, Taegtmeyer H (Dec 2014). "Targeting the ubiquitin-proteasome system in heart disease: the basis for new therapeutic strategies". Antioxidants & Redox Signaling. 21 (17): 2322–43. doi:10.1089/ars.2013.5823. PMC 4241867. PMID 25133688.
- Wang ZV, Hill JA (Feb 2015). "Protein quality control and metabolism: bidirectional control in the heart". Cell Metabolism. 21 (2): 215–26. doi:10.1016/j.cmet.2015.01.016. PMC 4317573. PMID 25651176.
- Karin M, Delhase M (Feb 2000). "The I kappa B kinase (IKK) and NF-kappa B: key elements of proinflammatory signalling". Seminars in Immunology. 12 (1): 85–98. doi:10.1006/smim.2000.0210. PMID 10723801.
- Ermolaeva MA, Dakhovnik A, Schumacher B (Jan 2015). "Quality control mechanisms in cellular and systemic DNA damage responses". Ageing Research Reviews. 23 (Pt A): 3–11. doi:10.1016/j.arr.2014.12.009. PMC 4886828. PMID 25560147.
- Checler F, da Costa CA, Ancolio K, Chevallier N, Lopez-Perez E, Marambaud P (Jul 2000). "Role of the proteasome in Alzheimer's disease". Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease. 1502 (1): 133–8. doi:10.1016/s0925-4439(00)00039-9. PMID 10899438.
- Chung KK, Dawson VL, Dawson TM (Nov 2001). "The role of the ubiquitin-proteasomal pathway in Parkinson's disease and other neurodegenerative disorders". Trends in Neurosciences. 24 (11 Suppl): S7–14. doi:10.1016/s0166-2236(00)01998-6. PMID 11881748.
- Ikeda K, Akiyama H, Arai T, Ueno H, Tsuchiya K, Kosaka K (Jul 2002). "Morphometrical reappraisal of motor neuron system of Pick's disease and amyotrophic lateral sclerosis with dementia". Acta Neuropathologica. 104 (1): 21–8. doi:10.1007/s00401-001-0513-5. PMID 12070660.
- Manaka H, Kato T, Kurita K, Katagiri T, Shikama Y, Kujirai K, Kawanami T, Suzuki Y, Nihei K, Sasaki H (May 1992). "Marked increase in cerebrospinal fluid ubiquitin in Creutzfeldt–Jakob disease". Neuroscience Letters. 139 (1): 47–9. doi:10.1016/0304-3940(92)90854-z. PMID 1328965.
- Mathews KD, Moore SA (Jan 2003). "Limb-girdle muscular dystrophy". Current Neurology and Neuroscience Reports. 3 (1): 78–85. doi:10.1007/s11910-003-0042-9. PMID 12507416.
- Mayer RJ (Mar 2003). "From neurodegeneration to neurohomeostasis: the role of ubiquitin". Drug News & Perspectives. 16 (2): 103–8. doi:10.1358/dnp.2003.16.2.829327. PMID 12792671.
- Calise J, Powell SR (Feb 2013). "The ubiquitin proteasome system and myocardial ischemia". American Journal of Physiology. Heart and Circulatory Physiology. 304 (3): H337–49. doi:10.1152/ajpheart.00604.2012. PMC 3774499. PMID 23220331.
- Predmore JM, Wang P, Davis F, Bartolone S, Westfall MV, Dyke DB, Pagani F, Powell SR, Day SM (Mar 2010). "Ubiquitin proteasome dysfunction in human hypertrophic and dilated cardiomyopathies". Circulation. 121 (8): 997–1004. doi:10.1161/CIRCULATIONAHA.109.904557. PMC 2857348. PMID 20159828.
- Powell SR (Jul 2006). "The ubiquitin–proteasome system in cardiac physiology and pathology" (PDF). American Journal of Physiology. Heart and Circulatory Physiology. 291 (1): H1–H19. doi:10.1152/ajpheart.00062.2006. PMID 16501026.
- Adams J (Apr 2003). "Potential for proteasome inhibition in the treatment of cancer". Drug Discovery Today. 8 (7): 307–15. doi:10.1016/s1359-6446(03)02647-3. PMID 12654543.
- Ben-Neriah Y (Jan 2002). "Regulatory functions of ubiquitination in the immune system". Nature Immunology. 3 (1): 20–6. doi:10.1038/ni0102-20. PMID 11753406.
- Egerer K, Kuckelkorn U, Rudolph PE, Rückert JC, Dörner T, Burmester GR, Kloetzel PM, Feist E (Oct 2002). "Circulating proteasomes are markers of cell damage and immunologic activity in autoimmune diseases". The Journal of Rheumatology. 29 (10): 2045–52. PMID 12375310.
- Matsuyama Y, Suzuki M, Arima C, Huang QM, Tomida S, Takeuchi T, Sugiyama R, Itoh Y, Yatabe Y, Goto H, Takahashi T (Apr 2011). "Proteasomal non-catalytic subunit PSMD2 as a potential therapeutic target in association with various clinicopathologic features in lung adenocarcinomas". Molecular Carcinogenesis. 50 (4): 301–9. doi:10.1002/mc.20632. PMID 21465578.
- Boldin MP, Mett IL, Wallach D (June 1995). "A protein related to a proteasomal subunit binds to the intracellular domain of the p55 TNF receptor upstream to its 'death domain'". FEBS Lett. 367 (1): 39–44. doi:10.1016/0014-5793(95)00534-G. PMID 7601280.
- Dunbar JD, Song HY, Guo D, Wu LW, Donner DB (May 1997). "Two-hybrid cloning of a gene encoding TNF receptor-associated protein 2, a protein that interacts with the intracellular domain of the type 1 TNF receptor: identity with subunit 2 of the 26S protease". J. Immunol. 158 (9): 4252–9. PMID 9126987.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (October 2005). "Towards a proteome-scale map of the human protein–protein interaction network". Nature. 437 (7062): 1173–8. Bibcode:2005Natur.437.1173R. doi:10.1038/nature04209. PMID 16189514.
- Gorbea C, Taillandier D, Rechsteiner M (January 2000). "Mapping subunit contacts in the regulatory complex of the 26 S proteasome. S2 and S5b form a tetramer with ATPase subunits S4 and S7". J. Biol. Chem. 275 (2): 875–82. doi:10.1074/jbc.275.2.875. PMID 10625621.
Further reading
- Coux O, Tanaka K, Goldberg AL (1996). "Structure and functions of the 20S and 26S proteasomes". Annu. Rev. Biochem. 65: 801–47. doi:10.1146/annurev.bi.65.070196.004101. PMID 8811196.
- Goff SP (2003). "Death by deamination: a novel host restriction system for HIV-1". Cell. 114 (3): 281–3. doi:10.1016/S0092-8674(03)00602-0. PMID 12914693.
- Boldin MP, Mett IL, Wallach D (1995). "A protein related to a proteasomal subunit binds to the intracellular domain of the p55 TNF receptor upstream to its 'death domain'". FEBS Lett. 367 (1): 39–44. doi:10.1016/0014-5793(95)00534-G. PMID 7601280.
- Song HY, Donner DB (1995). "Association of a RING finger protein with the cytoplasmic domain of the human type-2 tumour necrosis factor receptor". Biochem. J. 309 (3): 825–9. doi:10.1042/bj3090825. PMC 1135706. PMID 7639698.
- Seeger M, Ferrell K, Frank R, Dubiel W (1997). "HIV-1 tat inhibits the 20 S proteasome and its 11 S regulator-mediated activation". J. Biol. Chem. 272 (13): 8145–8. doi:10.1074/jbc.272.13.8145. PMID 9079628.
- Dunbar JD, Song HY, Guo D, Wu LW, Donner DB (1997). "Two-hybrid cloning of a gene encoding TNF receptor-associated protein 2, a protein that interacts with the intracellular domain of the type 1 TNF receptor: identity with subunit 2 of the 26S protease". J. Immunol. 158 (9): 4252–9. PMID 9126987.
- Madani N, Kabat D (1998). "An endogenous inhibitor of human immunodeficiency virus in human lymphocytes is overcome by the viral Vif protein". J. Virol. 72 (12): 10251–5. doi:10.1128/JVI.72.12.10251-10255.1998. PMC 110608. PMID 9811770.
- Simon JH, Gaddis NC, Fouchier RA, Malim MH (1998). "Evidence for a newly discovered cellular anti-HIV-1 phenotype". Nat. Med. 4 (12): 1397–400. doi:10.1038/3987. PMID 9846577.
- Gorbea C, Taillandier D, Rechsteiner M (2000). "Mapping subunit contacts in the regulatory complex of the 26 S proteasome. S2 and S5b form a tetramer with ATPase subunits S4 and S7". J. Biol. Chem. 275 (2): 875–82. doi:10.1074/jbc.275.2.875. PMID 10625621.
- Mulder LC, Muesing MA (2000). "Degradation of HIV-1 integrase by the N-end rule pathway". J. Biol. Chem. 275 (38): 29749–53. doi:10.1074/jbc.M004670200. PMID 10893419.
- You J, Pickart CM (2001). "A HECT domain E3 enzyme assembles novel polyubiquitin chains". J. Biol. Chem. 276 (23): 19871–8. doi:10.1074/jbc.M100034200. PMID 11278995.
- Sheehy AM, Gaddis NC, Choi JD, Malim MH (2002). "Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein". Nature. 418 (6898): 646–50. Bibcode:2002Natur.418..646S. doi:10.1038/nature00939. PMID 12167863.
- Huang X, Seifert U, Salzmann U, Henklein P, Preissner R, Henke W, Sijts AJ, Kloetzel PM, Dubiel W (2002). "The RTP site shared by the HIV-1 Tat protein and the 11S regulator subunit alpha is crucial for their effects on proteasome function including antigen processing". J. Mol. Biol. 323 (4): 771–82. doi:10.1016/S0022-2836(02)00998-1. PMID 12419264.
- You J, Wang M, Aoki T, Tamura TA, Pickart CM (2003). "Proteolytic targeting of transcriptional regulator TIP120B by a HECT domain E3 ligase". J. Biol. Chem. 278 (26): 23369–75. doi:10.1074/jbc.M212887200. PMID 12692129.
- Gaddis NC, Chertova E, Sheehy AM, Henderson LE, Malim MH (2003). "Comprehensive investigation of the molecular defect in vif-deficient human immunodeficiency virus type 1 virions". J. Virol. 77 (10): 5810–20. doi:10.1128/JVI.77.10.5810-5820.2003. PMC 154025. PMID 12719574.
- Lecossier D, Bouchonnet F, Clavel F, Hance AJ (2003). "Hypermutation of HIV-1 DNA in the absence of the Vif protein". Science. 300 (5622): 1112. doi:10.1126/science.1083338. PMID 12750511.
- Zhang H, Yang B, Pomerantz RJ, Zhang C, Arunachalam SC, Gao L (2003). "The cytidine deaminase CEM15 induces hypermutation in newly synthesized HIV-1 DNA". Nature. 424 (6944): 94–8. Bibcode:2003Natur.424...94Z. doi:10.1038/nature01707. PMC 1350966. PMID 12808465.