Ostracoderm
Ostracoderms (Ancient Greek, ὄστρακον+δέρμα "shell-skinned") are the armored jawless fish of the Paleozoic. The term does not often appear in classifications today because it is paraphyletic or polyphyletic, and thus does not correspond to one evolutionary lineage.[1] However, the term is still used as an informal way of loosely grouping together the armored jawless fishes.
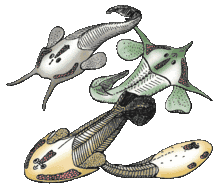

An innovation of ostracoderms was the use of gills not for feeding, but exclusively for respiration. Earlier chordates with gills used them for both respiration and feeding. Ostracoderms had separate pharyngeal gill pouches along the side of the head, which were permanently open with no protective operculum. Unlike invertebrates that use ciliated motion to move food, ostracoderms used their muscular pharynx to create a suction that pulled small and slow moving prey into their mouths.
The first fossil fishes that were discovered were ostracoderms. The Swiss anatomist Louis Agassiz received some fossils of bony armored fish from Scotland in the 1830s. He had a hard time classifying them as they did not resemble any living creature. He compared them at first with extant armored fish such as catfish and sturgeons but later realizing that they had no movable jaws, classified them in 1844 into a new group "ostracoderms" which means "shell-skinned".[2]
Ostracoderms have heads covered with a bony shield. They are among the earliest creatures with bony heads. The microscopic layers of that shield appear to evolutionary biologists, "like they are composed of little tooth-like structures."[3] Neil Shubin writes: "Cut the bone of the [ostracoderm] skull open…pop it under a microscope and…you find virtually the same structure as in our teeth. There is a layer of enamel and even a layer of pulp. The whole shield is made up of thousands of small teeth fused together. This bony skull--one of the earliest in the fossil record--is made entirely of little teeth. Teeth originally arose to bite creatures (see Conodonts); later a version of teeth was used in a new way to protect them."[3]
Ostracoderms existed in two major groups, the more primitive heterostracans and the cephalaspids. The cephalaspids were more advanced than the heterostracans in that they had lateral stabilizers for more control of their swimming.
It was long assumed that pteraspidomorphs and thelodonts were the only ostracoderms with paired nostrils, while the other groups have just a single median nostril. But it has since been revealed that even if galeaspidans have just one external opening, it has two internal nasal organs.[4][5]
After the appearance of jawed fish (placoderms, acanthodians, sharks, etc.) about 420 million years ago, most ostracoderm species underwent a decline, and the last ostracoderms became extinct at the end of the Devonian period. More recent research indicates, however, that fish with jaws had far less to do with the extinction of the ostracoderms than previously assumed, as they coexisted without noticeable decline for about 30 million years.[6]
The Subclass Ostracodermi has been placed in the division Agnatha along with the extant Subclass Cyclostomata, which includes lampreys and hagfishes.
Major groups
| Major groups of ostracoderms | |||
|---|---|---|---|
| Group | Class | Image | Description |
| Cephalaspido- morphi |
Cephalaspidomorphi, or cephalaspids, were like most contemporary fishes very well armoured. Particularly the head shield was well developed, protecting the head, gills and the anterior section of innards. The body were in most forms well armoured too. The head shield had a series of grooves over the whole surface forming an extensive lateral line organ. The eyes were rather small and placed on the top of the head. There was no jaw proper. The mouth opening was surrounded by small plates making the lips flexible, but without any ability to bite.[7] Most biologists regard this taxon as extinct, but the name is sometimes used in the classification of lampreys because lampreys were once thought to be related to cephalaspids. If lampreys are included, they would extend the known range of the group from the Silurian and Devonian periods to the present day. | ||
| Galeaspida (extinct) |
 |
Galeaspida (helmet shields) have massive bone shield on the head. Galeaspida lived in shallow, fresh water and marine environments during the Silurian and Devonian times (430 to 370 million years ago) in what is now Southern China, Tibet and Vietnam. Superficially, their morphology appears more similar to that of Heterostraci than Osteostraci, there being currently no evidence that the galeaspids had paired fins. However, Galeaspida are in fact regarded as being more closely related to Osteostraci, based on the closer similarity of the morphology of the braincase. | |
| Pituriaspida (extinct) |
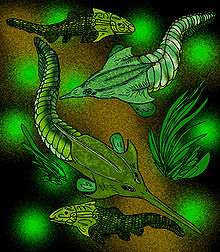 |
Pituriaspida (pituri or hallucinogenic shield) are a small group of extinct armoured jawless fishes with tremendous nose-like rostrums, which lived in the marine, deltaic environments of Middle Devonian Australia (about 390 Ma). They are known only by two species, Pituriaspis doylei and Neeyambaspis enigmatica found in a single sandstone location of the Georgina Basin, in Western Queensland, Australia | |
| Osteostraci (extinct) |
|
Osteostraci (bony shields) lived in what is now North America, Europe and Russia from the Middle Silurian to Late Devonian. Anatomically speaking, the osteostracans, especially the Devonian species, were among the most advanced of all known agnathans. This is due to the development of paired fins, and their complicated cranial anatomy. The osteostracans were more similar to lampreys than to jawed vertebrates in possessing two pairs of semicircular canals in the inner ear, as opposed to the three pairs found in the inner ears of jawed vertebrates. They are thought to be the sister-group of pituriaspids, and together, these two taxa of jawless vertebrates are the sister-group of gnathostomes. Several synapomorphies support this hypothesis, such as the presence of: sclerotic ossicles, paired pectoral fins, a dermal skeleton with three layers (a basal layer of isopedin, a middle layer of spongy bone, and a superficial layer of dentin), and perichondral bone.[8] | |
| Other groups |
Other groups | ||
| †Pteraspido- morphi (extinct) |
 |
†Pteraspidomorphi have extensive shielding of the head. Many had hypocercal tails in order to generate lift to increase ease of movement through the water for their armoured bodies, which were covered in dermal bone. They also had sucking mouth parts and some species may have lived in fresh water.
The taxon contains the subgroups Heterostraci, Astraspida, Arandaspida. | |
| †Thelodonti (extinct) |
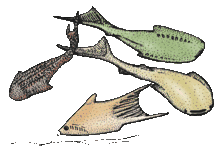 |
Thelodonti (nipple teeth) are a group of small, extinct jawless fishes with distinctive scales instead of large plates of armour. There is much debate over whether the group of Palaeozoic fish known as the Thelodonti (formerly coelolepids[9]) represent a monophyletic grouping, or disparate stem groups to the major lines of jawless and jawed fish. Thelodonts are united in possession of "thelodont scales". This defining character is not necessarily a result of shared ancestry, as it may have been evolved independently by different groups. Thus the thelodonts are generally thought to represent a polyphyletic group,[10] although there is no firm agreement on this point; if they are monophyletic, there is no firm evidence on what their ancestral state was.[11]:206 "Thelodonts" were morphologically very similar, and probably closely related, to fish of the classes Heterostraci and Anaspida, differing mainly in their covering of distinctive, small, spiny scales. These scales were easily dispersed after death; their small size and resilience makes them the most common vertebrate fossil of their time.[12][13] The fish lived in both freshwater and marine environments, first appearing during the Ordovician, and perishing during the Frasnian–Famennian extinction event of the Late Devonian. They were predominantly deposit-feeding bottom dwellers, although there is evidence to suggest that some species took to the water column to be free-swimming organisms. | |
| †Anaspida (extinct) |
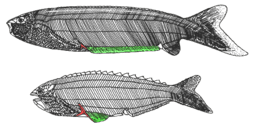 |
Anaspida (without shield) is an extinct group of primitive jawless vertebrates that lived during the Silurian and Devonian periods.[14] They are classically regarded as the ancestors of lampreys.[15] Anaspids were small marine agnathans that lacked heavy bony shield and paired fins, but have a striking highly hypocercal tail. They first appeared in the Early Silurian, and flourished until the Late Devonian extinction,[16] where most species, save for lampreys, became extinct due to the environmental upheaval during that time. | |
See also
References
- Benton, Michael (2009) Vertebrate Palaeontology Edition 3, page 44, John Wiley & Sons. ISBN 9781405144490.
- Maisey, John G. (1996). Discovering Fossil Fishes (illustrated ed.). New York: Henry Holt & Company. p. 37.
- Shubin, Neil (2009). Your Inner Fish: A Journay into the 3.5 Billion Year History of the Human Body (reprint ed.). New York: Pantheon Books. pp. 85–86. ISBN 9780307277459.
- "Fossil fish reveals how jaws evolved - Planet Earth Online". Archived from the original on 2012-07-03. Retrieved 2014-04-11.
- GEOL 331 Invertebrate Paleontology
- Vertebrate jaw design locked down early
- Morales, Edwin H. Colbert, Michael (1991). Evolution of the vertebrates : a history of the backboned animals through time (4th ed.). New York: Wiley-Liss. ISBN 978-0-471-85074-8.
- Sansom, R. S. (2009). "Phylogeny, classification and character polarity of the Osteostraci (Vertebrata)". Journal of Systematic Palaeontology. 7: 95–115. doi:10.1017/S1477201908002551.
- Turner, S.; Tarling, D. H. (1982). "Thelodont and other agnathan distributions as tests of Lower Paleozoic continental reconstructions". Palaeogeography, Palaeoclimatology, Palaeoecology. 39 (3–4): 295–311. doi:10.1016/0031-0182(82)90027-X.
- Sarjeant, William Antony S.; L. B. Halstead (1995). Vertebrate fossils and the evolution of scientific concepts: writings in tribute to Beverly Halstead. ISBN 978-2-88124-996-9.
- Donoghue, P. C., P. L. Forey & R. J. Aldridge (2000). "Conodont affinity and chordate phylogeny". Biological Reviews of the Cambridge Philosophical Society. 75 (2): 191–251. doi:10.1111/j.1469-185X.1999.tb00045.x. PMID 10881388.CS1 maint: multiple names: authors list (link)
- Turner, S. (1999). "Early Silurian to Early Devonian thelodont assemblages and their possible ecological significance". In A. J. Boucot; J. Lawson (eds.). Palaeocommunities, International Geological Correlation Programme 53, Project Ecostratigraphy, Final Report. Cambridge University Press. pp. 42–78.
- The early and mid Silurian. See Kazlev, M.A., White, T (March 6, 2001). "Thelodonti". Palaeos.com. Archived from the original on October 28, 2007. Retrieved October 30, 2007.CS1 maint: multiple names: authors list (link)
- Ahlberg, Per Erik (2001). Major events in early vertebrate evolution: palaeontology, phylogeny, genetics, and development. Washington, DC: Taylor & Francis. p. 188. ISBN 0-415-23370-4.
- Patterson, Colin (1987). Molecules and morphology in evolution: conflict or compromise?. Cambridge, UK: Cambridge University Press. p. 142. ISBN 0-521-32271-5.
- Hall, Brian Keith; Hanken, James (1993). The Skull. Chicago: University of Chicago Press. p. 131. ISBN 0-226-31568-1.
| Wikispecies has information related to Ostracodermi |
| Forerunners |  | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jawless fish |
| ||||||||||||
| Jawed fish |
| ||||||||||||
| Lists | |||||||||||||
| Related |
| ||||||||||||
† extinct | |||||||||||||