Leghemoglobin reductase
In enzymology, a leghemoglobin reductase (EC 1.6.2.6) is an enzyme that catalyzes the chemical reaction
- NAD(P)H + H+ + 2 ferrileghemoglobin NAD(P)+ + 2 ferroleghemoglobin

| leghemoglobin reductase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 1.6.2.6 | ||||||||
| CAS number | 60440-35-9 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / QuickGO | ||||||||
| |||||||||
The 4 substrates of this enzyme are NADH, NADPH, H+, and ferrileghemoglobin, whereas its 3 products are NAD+, NADP+, and ferroleghemoglobin.
This enzyme belongs to the family of oxidoreductases, specifically those acting on NADH or NADPH with a heme protein as acceptor. The systematic name of this enzyme class is NAD(P)H:ferrileghemoglobin oxidoreductase. This enzyme is also called ferric leghemoglobin reductase.
Role of ferrous leghemoglobin in legume nodules.
Leghemoglobin (Lb) is a heme-containing protein that reversibly binds and transports O2 into the N2-fixing nodules of leguminous plants.[1] In order to function as an O2-carrier Lb must be in the ferrous oxidation state (Lb2+). Oxygenated Lb2+ (Lb2+O2) readily autoxidizes to ferric Lb (Lb3+) generating O2− in the presence of trace amounts of transition metals, chelators and toxic metabolites (such as nitrite, superoxide radical and peroxides),[2] however Lb2+ is the predominant form in nodules.[3][4] Therefore, mechanisms exist in vivo for maintaining Lb in the functional ferrous status.[5]
The discovery of ferric leghemoglobin reductases.
Burris and Hass[6] were the first to propose that reduced pyridine nucleotides might function as reductants of Lb3+ in leguminous root nodules and in 1969 Appleby[7] reported that Lb3+ was reduced to Lb2+ by a suspension of bacteroids. In 1982 Kretovich and collaborators[8] purified an enzyme from lupine nodules which catalyzed the reduction of Lb3+ to Lb2+ using NADH as reductant. This enzyme (named by these authors as Legoglobin Reductase -LR) is similar to NADH:cytochrome b5 reductase (EC 1.6.2.2) from erythrocytes and bovine muscle. Lupin LR is a flavoprotein with a molecular mass of 60 kDa and its activity is specific for NADH. In 1984 Klucas and collaborators[9] purified a protein with ferric Lb reductase (FLbR) activity from soybean nodules. The activity of soybean FLbR was 90% in the nodule cytosol and 10% in the bacteroids. NADH was the best reductant for soybean FLbR, although NADPH also functioned at rates that were three-fold less than NADH. These investigations by Klucas and collaborators[9] also showed that the oxidation of NADH and reduction of Lb3+ was undetectable when O2 was removed from the reaction system, but all were restored upon re-addition of O2, which indicated that the FLbR activity is O2-dependent.
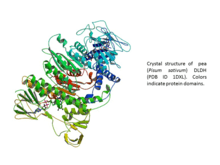
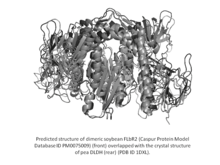
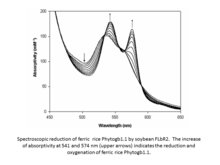
Soybean and other legume ferric leghemoglobin reductases.
Soybean FLbR is a flavoprotein with flavin adenine dinucleotide (FAD) as the prosthetic group and consists of two identical subunits, each having a molecular mass of 54 kDa. The Km and Kcat values of soybean FLbR for soybean Lb3+ reduction are 9.2 μM and 6.2 s−1, respectively (Kcat/Km = 674 M−1 s−1). The amino acid sequence of soybean FLbR is highly related to that of the flavin-nucleotide disulfide oxidoreductases, especially dihydrolipoamide dehydrogenase (DLDH) (EC 1.8.1.4) of the pyruvate dehydrogenase complex. The amino acid sequence of soybean FLbR contains a 30-residue signal peptide for translocation into the mitochondria as well as conserved regions for the FAD-binding site, NAD(P)H-binding site and disulfide active site characteristic of pea DLDH and other enzymes in the family of the pyridine nucleotide-disulfide oxidoreductases.[10]
The soybean genome contains at least two copies (named flbr1 and flbr2) of the flbr gene.[11] The amino acid sequence of soybean FLbR2 has considerable homology with soybean FLbR1 and pea leaf mitochondria DLDH and contains a 30-residue mitochondrial transit peptide.[12] FLbR sequences have also been detected and analyzed in legumes other than soybean. For example, the nucleotide sequence of a cowpea FLbR cDNA has 88 and 85% similarity with soybean FLbR and pea DLDH, respectively. The Km and Kcat values of cowpea FLbR for cowpea Lb3+ reduction are 10.4 μM and 3.1 s−1, respectively (Kcat/Km = 298 M−1 s−1).[13]
Ferric leghemoglobin reductases as possible reductants of phytoglobins in vivo.
Soybean FLbR2 reduces ferric rice Phytoglobin1.1 (Phytogb1.13+).[14] Apparently, the soybean FLbR2-rice Phytoglobin1.13+ interaction is weak. An in silico analysis predicted that soybean FLbR2 and rice Phytogb1.13+ interact at the FAD-binding domain of soybean FLbR2 and the CD-loop and helix F of rice Phytogb1.13+. Therefore, FLbRs could be a generalized in vivo mechanism for the enzymatic reduction of Phytogbs3+.
References
- Appleby C. A., The origin and functions of haemoglobin in plants, Sci. Progress, 76 (1992) 365-398.
- Becana M.,Klucas R. V., Oxidation and reduction of leghemoglobin in root nodules of leguminous plants, Plant Physiol., 98 (1992) 1217-1221.
- Lee K. K.,Klucas R. V., Reduction of ferric leghemoglobin in soybean root nodules, Plant Physiol., 74 (1984) 984-988.
- Lee K. K., Shearman L. L., Ericson B. K.,Klucas R. V., Ferric leghemoglobin in plant-attached leguminous nodules., Plant Physiol., 109 (1995) 261-267.
- Becana M.,Klucas R. V., Enzymatic and nonenzymatic mechanisms for ferric leghemoglobin reduction in legume root nodules, Proc. Natl. Acad. Sci. USA, 87 (1990) 7295-7299.
- Burris R. H.,Hass E., The red pigment of leguminous root nodules., J. Biol. Chem., 155 (1944) 227-229.
- Appleby C. A., Properties of leghaemoglobin in vivo, and its isolation as ferrous oxyleghaemoglobin, Biochim Biophys. Acta, 188 (1969) 222-229.
- Kretovich V. L., Melik-Sarkisyan S. S., Bashirova N. F.,Topunov A. F., Enzymatic reduction of leghemoglobin in lupin nodules, J. Appl. Biochem., 4 (1982) 209-217.
- Saari L. L.,Klucas R. V., Ferric leghemoglobin reductase from soybean root nodules, Arch. Biochem. Biophys., 231 (1984) 102-113.
- Ji L., Wood S., Becana M.,Klucas R. V., Purification and characterization of soybean root nodule ferric leghemoglobin reductase, Plant Physiol., 96 (1991) 32-37.
- Ji L., Becana M., Sarath G.,Klucas R. V., Cloning and sequence analysis of a cDNA encoding ferric leghemoglobin reductase from soybean nodules, Plant Physiol., 104 (1994) 453-459.
- Moran J. F., Sun Z., Sarath G., Arredondo-Peter R., James E. K., Becana M.,Klucas R. V., Molecular cloning, functional characterization, and subcellular localization of soybean nodule dihydrolipoamide reductase., Plant Physiol., 128 (2002) 300-313.
- Luan P., Aréchaga-Ocampo E., Sarath G., Arredondo-Peter R.,Klucas R. V., Analysis of a ferric leghemoglobin reductase from cowpea (Vigna unguiculata) root nodules., Plant Sci., 154 (2000) 161-170.
- Gopalasubramaniam S. K., Kondapalli K. C., Millán-Pacheco C., Pastor N., Stemmler T. L., Moran J. F.,Arredondo-Peter R., Soybean dihydrolipoamide dehydrogenase (ferric leghemoglobin reductase 2) interacts with and reduces ferric non-symbiotic hemoglobin 1., ScienceJet, 2 (2013) 33.
- Saari LL, Klucas RV (1984). "Ferric leghemoglobin reductase from soybean root nodules". Arch. Biochem. Biophys. 231 (1): 102–13. doi:10.1016/0003-9861(84)90367-9. PMID 6539095.