Hopanoids
Hopanoids are a diverse subclass of triterpenoids with the same hydrocarbon skeleton as the compound hopane. This group of pentacyclic molecules therefore refers to simple hopenes, hopanols and hopanes, but also to extensively functionalized derivatives such as bacteriohopanepolyols (BHP) and hopanoids covalently attached to lipid A.[1][2]
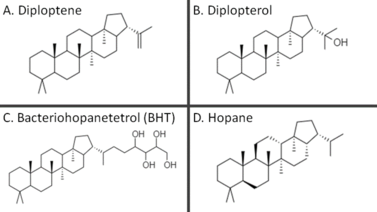
The first known hopanoid, hydroxyhopanone, was isolated by two chemists at The National Gallery, London working on the chemistry of dammar gum, a natural resin used as a varnish for paintings.[3] The name hopane was derived from the genus Hopea (a source of the resin), which in turn was named after the botanist John Hope. Since this first finding in an angiosperm, however, hopanoids have been found in membranes of bacteria, lichens, bryophytes, ferns and fungi.[4] Because of their geostable structures, various hopanoids are preserved in petroleum reservoirs, rocks and sediment, allowing them to be used as biological markers.[5] Hopanoids have not been detected in archaea.[6][7]
Biological function
About 10% of sequenced bacterial genomes have a putative shc gene encoding a squalene-hopene cyclase and can presumably make hopanoids, which have been shown to play diverse roles in the plasma membrane and may allow some organisms to adapt in extreme environments.[8][8]
Since hopanoids modify plasma membrane properties in bacteria, they are frequently compared to sterols, such as cholesterol, which modulate membrane fluidity and serve other functions in eukaryotes.[9] Although hopanoids do not seem to rescue sterol deficiency, they do condense membranes and decrease permeability.[10] Also, gammaproteobacteria and eukaryotic organisms such as lichens and bryophytes have been shown to produce both sterols and hopanoids, suggesting these lipids may have other distinct functions.[4][11] Notably, the way hopanoids pack into the plasma membrane can change depending on what functional groups are attached. The hopanoid bacteriohopanetetrol assumes a transverse orientation in lipid bilayers, but diploptene localizes between the inner and outer leaflet, presumably thickening the membrane to decrease permeability.[12]
The hopanoid diplopterol orders membranes by interacting with lipid A, a common membrane lipid in bacteria, in ways similar to how cholesterol and sphingolipids interact in eukaryotic plasma membranes.[9] Diplopterol and cholesterol were demonstrated to promote condensation and inhibit gel-phase formation in both sphingomyelin monolayers and monolayers of glycan-modified lipid A. Furthermore, both diplopterol and cholesterol could rescue pH-dependent phase transitions in glycan-modified lipid A monolayers.[9] The role of hopanoids in membrane-mediated acid tolerance is further supported by observations of acid-inhibited growth and morphological abnormalities of the plasma membrane in hopanoid-deficient bacteria with mutant squalene-hopene cyclases.[13][14]
Hopanoids are produced in several nitrogen-fixing bacteria.[10] In the actinomycete Frankia, hopanoids in diazovesicle membranes likely restrict the entry of oxygen by making the lipid bilayer more tight and compact.[15] In Bradyrhizobium, hopanoids chemically bonded to lipid A increase membrane stability and rigidity, enhancing stress tolerance and intracellular survival in Aeschynomene legumes.[16] In the cyanobacterium Nostoc punctiforme, large quantities of 2-methylhopanoids localize to the outer membranes of survival structures called akinetes.[17] In another example of stress tolerance, hopanoids in the aerial hyphae (spore bearing structures) of the prokaryotic soil bacteria Streptomyces are thought to minimize water loss across the membrane to the air.[18]
Biosynthesis
Squalene synthesis
Since hopanoids are a C₃₀ terpenoid, biosynthesis begins with isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAP), which are combined to form longer chain isoprenoids.[2] Synthesis of these smaller precursors proceeds either via the mevalonate pathway or the methylerythritol-4-phosphate pathway depending on the bacterial species, although the latter tends to be more common.[19] DMAP condenses with one molecule of IPP to geranyl pyrophosphate, which in turn condenses with another IPP to generate farnesyl pyrophosphate.[2] Squalene synthase, coded for by the gene sqs, then catalyzes the condensation of two farnesyl pyrophosphate molecules to squalene while oxidizing NADPH.[20]
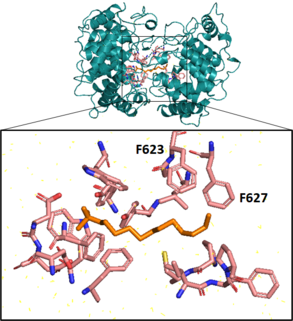
Cyclization
Next, a squalene-hopene cyclase catalyzes an elaborate cyclization reaction, engaging squalene in an energetically favorable all-chair conformation before creating 5 cycles, 6 covalent bonds, and 9 chiral centers on the molecule in a single step.[21][22] This enzyme, coded for by the gene shc, has a double ⍺-barrel fold characteristic of terpenoid biosynthesis[23] and is present in the cell as a monotopic homodimer, meaning pairs of the cyclase are embedded in but do not span the plasma membrane.[21][24] In vitro, this enzyme exhibits promiscuous substrate specificity, also cyclizing 2,3-oxidosqualene.[25]
Aromatic residues in the active site form several unfavorable carbocations on the substrate which are quenched by a rapid polycyclization.[22] In the last substep of the cyclization reaction, after electrons comprising the terminal alkene bond on the squalene have attacked the hopenyl carbocation to close the E ring, the C-22 carbocation may be quenched by mechanisms that lead to different hopanoid products. Nucleophilic attack of water will yield diplopterol, while deprotonation at an adjacent carbon will form one of several hopene isomers, often diploptene.[4]
Functionalization
After cyclization, hopanoids are frequently modified by hopanoid biosynthesis enzymes encoded by genes in the same operon as shc, hpn.[26] For instance, the radical SAM protein HpnH adds an adenosine group to diploptene, forming the extended C35 hopanoid adenosylhopane, which can then be further modified by other hpn gene products to form bacteriohopanetetrol (BHT).[27][28] Additional modification occurs as the glycosyltransferase HpnI converts BHT to N-acetylglucosaminyl-BHT.[29] In sequence, the hopanoid biosynthesis associated protein HpnK mediates deacetylation to glucosaminyl-BHT, from which radical SAM protein HpnJ generates a cyclitol ether.[29]
Importantly, C30 and C35 hopanoids alike may be methylated at C-2 and C-3 positions by the radical SAM methyltransferases HpnP and HpnR, respectively.[30][31] These two methylations are particularly geostable compared to side-chain modifications and have entertained geobiologists for decades.[10]
In a biosynthetic pathway exclusive to some bacteria, diploptene is a precursor for the pentacyclic triterpenoid tetrahymanol, which instead is produced directly from squalene by a cyclase in eukaryotes.[32]
In paleobiology
Hopanoids have been estimated to be the most abundant natural products on Earth, remaining in the organic fraction of all sediments, independent of age, origin or nature.[33] Biomolecules like DNA and proteins are degraded during diagenesis, but polycyclic lipids persist in the environment over geologic timescales due to their fused, stable structures.[34] Although hopanoids and sterols are reduced to hopanes and steranes during deposition, these diagenetic products can still be useful biomarkers, or molecular fossils, for studying the coevolution of early life and Earth.[34][35]
2-methylhopanes supposedly from photosynthetic cyanobacteria and steranes were reported by Roger Summons and colleagues as molecular fossils preserved in 2.7 Gya shales from the Pilbara region in Western Australia.[36] The presence of abundant 2-alpha-methylhopanes preserved in these shales was interpreted as evidence of oxygenic photosynthesis at least 2.7 Gya, unexpectedly suggesting a 400 million year gap between the evolution of oxygenic metabolism and when atmosphere became oxidizing. This interpretation of the biomarker record was challenged when Geobacter sulfurreducens was demonstrated to synthesize diverse hopanols, although not 2-methyl-hopanols, when grown under strictly anaerobic conditions.[8] The integrity of 2-methylhopanes as biomarkers for oxygenic photosynthesis was then further attenuated by evidence that the phototroph Rhodopseudomonas palustris produced 2-methyl-BHPs only under anoxic conditions.[37] Concrete evidence came from studies showing that not all cyanobacteria make hopanoids and that genes coding for the methyltransferase HpnP are present in nonphotosynthetic alphaproteobacteria and acidobacteria.[30][38]
Biomarker findings in the Pilbara-Craton shales were later rejected altogether during more recent evaluations of the fossil triterpenoid record.[39] Currently, the oldest detected triterpenoid fossils are Mesoproterozoic okenanes, steranes, and methylhopanes from a 1.64 Gya basin in Australia.[40] However, molecular clock analyses estimate that the earliest sterols were likely produced around 2.3 Gya, around the same time as the Great Oxidation Event, with hopanoid synthesis arising even earlier.[41]
For several reasons, hopanoids and squalene-hopene cyclases have been hypothesized to be more ancient than sterols and oxidosqualene cyclases. First, diplopterol is formed from water quenching the C-22 carbocation, meaning it can be made without molecular oxygen and could have served as a sterol surrogate before Earth's atmosphere became oxic. [1] Furthermore, squalene binds to squalene-hopene cyclases in a low-energy, all-chair conformation while oxidosqualene is cyclized in a more strained, chair-boat-chair-boat conformation.[4][42] Squalene-hopene cyclases also display more substrate promiscuity in that they cyclize oxidosqualene in vitro, causing some scientists to hypothesize that they are evolutionary predecessors to oxidosqualene cyclases.[42] Other scientists have proposed that squalene-hopene and oxidosqualene cyclases diverged from a common ancestor, a putative bacterial cyclase that would have made a tricyclic malabaricanoid or tetracyclic dammarinoid product.[1][43]
Industrial Relevance
The elegant mechanism behind the protonase activity of squalene-hopene cyclase was appreciated and adapted by chemical engineers at the University of Stuttgart, Germany. Active site engineering resulted in loss of the enzyme's ability to form hopanoids, but enabled Brønsted acid catalysis for the stereoselective cyclization of the monoterpenoids geraniol, epoxygeraniol, and citronellal.[44]
Agricultural Relevance
The application of hopanoids and hopanoid-producing nitrogen fixers has been proposed and patented as a biofertilizer technique that increases environmental resistance of plant-associated microbial symbionts.[45]
Medical Relevance
During later studies of interactions between diplopterol and lipid A in Methylobacterium extorquens, multidrug transport was found to be a hopanoid-dependent process. Squalene-hopene cyclase mutants derived from a wild type capable of multidrug efflux, a drug-resistance mechanism mediated by integral transport proteins, lost the ability to perform both multidrug transport and hopanoid synthesis.[46] Researchers indicate this could be due to direct regulation of transport proteins by hopanoids or indirectly by altering membrane ordering in a way that disrupts the transport system.[46]
References
- Welander, Paula V. (2019-08-20). "Deciphering the evolutionary history of microbial cyclic triterpenoids". Free Radical Biology and Medicine. Early Life on Earth and Oxidative Stress. 140: 270–278. doi:10.1016/j.freeradbiomed.2019.05.002. ISSN 0891-5849. PMID 31071437.
- Sohlenkamp, Christian; Geiger, Otto (2016-01-01). "Bacterial membrane lipids: diversity in structures and pathways". FEMS Microbiology Reviews. 40 (1): 133–159. doi:10.1093/femsre/fuv008. ISSN 0168-6445. PMID 25862689.
- Mills, J. S.; Werner, A. E. A. (1955-01-01). "The chemistry of dammar resin". Journal of the Chemical Society (Resumed): 3132–3140. doi:10.1039/JR9550003132. ISSN 0368-1769.
- Volkman, John K. (2005-02-01). "Sterols and other triterpenoids: source specificity and evolution of biosynthetic pathways". Organic Geochemistry. 36 (2): 139–159. doi:10.1016/j.orggeochem.2004.06.013. ISSN 0146-6380.
- Hunt, John M.; Philp, R. Paul; Kvenvolden, Keith A. (2002-09-01). "Early developments in petroleum geochemistry". Organic Geochemistry. 33 (9): 1025–1052. doi:10.1016/S0146-6380(02)00056-6. ISSN 0146-6380.
- William W. Christie. "The AOCS Lipid Library. Hopanoids". American Oil Chemists' Society. Archived from the original on 2016-03-05. Retrieved 2015-11-17.
- "Hopanoids - AOCS Lipid Library". 2016-03-05. Archived from the original on 2016-03-05. Retrieved 2020-03-06.
- Fischer, W. W.; Summons, R. E.; Pearson, A. (2005). "Targeted genomic detection of biosynthetic pathways: anaerobic production of hopanoid biomarkers by a common sedimentary microbe". Geobiology. 3 (1): 33–40. doi:10.1111/j.1472-4669.2005.00041.x. ISSN 1472-4669.
- Sáenz, James Peter; Sezgin, Erdinc; Schwille, Petra; Simons, Kai (2012-08-28). "Functional convergence of hopanoids and sterols in membrane ordering". Proceedings of the National Academy of Sciences. 109 (35): 14236–14240. doi:10.1073/pnas.1212141109. ISSN 0027-8424. PMC 3435179. PMID 22893685.
- Belin, Brittany J.; Busset, Nicolas; Giraud, Eric; Molinaro, Antonio; Silipo, Alba; Newman, Dianne K. (May 2018). "Hopanoid lipids: from membranes to plant–bacteria interactions". Nature Reviews Microbiology. 16 (5): 304–315. doi:10.1038/nrmicro.2017.173. ISSN 1740-1534. PMC 6087623. PMID 29456243.
- Tippelt, Anette; Jahnke, Linda; Poralla, Karl (1998-03-30). "Squalene–hopene cyclase from Methylococcus capsulatus (Bath): a bacterium producing hopanoids and steroids". Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 1391 (2): 223–232. doi:10.1016/S0005-2760(97)00212-9. ISSN 0005-2760. PMID 9555026.
- Poger, David; Mark, Alan E. (2013-12-19). "The Relative Effect of Sterols and Hopanoids on Lipid Bilayers: When Comparable Is Not Identical". The Journal of Physical Chemistry B. 117 (50): 16129–16140. doi:10.1021/jp409748d. ISSN 1520-6106. PMID 24299489.
- Schmerk, Crystal L.; Bernards, Mark A.; Valvano, Miguel A. (2011-12-01). "Hopanoid Production Is Required for Low-pH Tolerance, Antimicrobial Resistance, and Motility in Burkholderia cenocepacia". Journal of Bacteriology. 193 (23): 6712–6723. doi:10.1128/JB.05979-11. ISSN 0021-9193. PMC 3232912. PMID 21965564.
- Welander, Paula V.; Hunter, Ryan C.; Zhang, Lichun; Sessions, Alex L.; Summons, Roger E.; Newman, Dianne K. (2009-10-01). "Hopanoids Play a Role in Membrane Integrity and pH Homeostasis in Rhodopseudomonas palustris TIE-1". Journal of Bacteriology. 191 (19): 6145–6156. doi:10.1128/JB.00460-09. ISSN 0021-9193. PMC 2747905. PMID 19592593.
- Berry, A. M.; Harriott, O. T.; Moreau, R. A.; Osman, S. F.; Benson, D. R.; Jones, A. D. (1993-07-01). "Hopanoid lipids compose the Frankia vesicle envelope, presumptive barrier of oxygen diffusion to nitrogenase". Proceedings of the National Academy of Sciences. 90 (13): 6091–6094. doi:10.1073/pnas.90.13.6091. ISSN 0027-8424. PMC 46873. PMID 11607408.
- Silipo, Alba; Vitiello, Giuseppe; Gully, Djamel; Sturiale, Luisa; Chaintreuil, Clémence; Fardoux, Joel; Gargani, Daniel; Lee, Hae-In; Kulkarni, Gargi; Busset, Nicolas; Marchetti, Roberta (2014-10-30). "Covalently linked hopanoid-lipid A improves outer-membrane resistance of a Bradyrhizobium symbiont of legumes". Nature Communications. 5 (1): 5106. doi:10.1038/ncomms6106. ISSN 2041-1723. PMID 25355435.
- Doughty, D. M.; Hunter, R. C.; Summons, R. E.; Newman, D. K. (2009). "2-Methylhopanoids are maximally produced in akinetes of Nostoc punctiforme: geobiological implications". Geobiology. 7 (5): 524–532. doi:10.1111/j.1472-4669.2009.00217.x. ISSN 1472-4669. PMC 2860729. PMID 19811542.
- Poralla, Karl; Muth, Günther; Härtner, Thomas (2000-08-01). "Hopanoids are formed during transition from substrate to aerial hyphae in Streptomyces coelicolor A3(2)". FEMS Microbiology Letters. 189 (1): 93–95. doi:10.1111/j.1574-6968.2000.tb09212.x. ISSN 0378-1097. PMID 10913872.
- Pérez-Gil, Jordi; Rodríguez-Concepción, Manuel (2013-05-15). "Metabolic plasticity for isoprenoid biosynthesis in bacteria". Biochemical Journal. 452 (1): 19–25. doi:10.1042/BJ20121899. ISSN 0264-6021. PMID 23614721.
- Pan, Jian-Jung; Solbiati, Jose O.; Ramamoorthy, Gurusankar; Hillerich, Brandan S.; Seidel, Ronald D.; Cronan, John E.; Almo, Steven C.; Poulter, C. Dale (2015-05-27). "Biosynthesis of Squalene from Farnesyl Diphosphate in Bacteria: Three Steps Catalyzed by Three Enzymes". ACS Central Science. 1 (2): 77–82. doi:10.1021/acscentsci.5b00115. ISSN 2374-7943. PMC 4527182. PMID 26258173.
- Siedenburg, Gabriele; Jendrossek, Dieter (2011-06-15). "Squalene-Hopene Cyclases". Applied and Environmental Microbiology. 77 (12): 3905–3915. doi:10.1128/AEM.00300-11. ISSN 0099-2240. PMC 3131620. PMID 21531832.
- Hoshino, Tsutomu; Sato, Tsutomu (2002-02-12). "Squalene–hopene cyclase: catalytic mechanism and substrate recognition". Chemical Communications (4): 291–301. doi:10.1039/B108995C. ISSN 1364-548X. PMID 12120044.
- Syrén, Per-Olof; Henche, Sabrina; Eichler, Anja; Nestl, Bettina M.; Hauer, Bernhard (2016-12-01). "Squalene-hopene cyclases—evolution, dynamics and catalytic scope". Current Opinion in Structural Biology. Multi-protein assemblies in signaling • Catalysis and regulation. 41: 73–82. doi:10.1016/j.sbi.2016.05.019. ISSN 0959-440X. PMID 27336183.
- Dang, Tongyun; Prestwich, Glenn D. (2000-08-01). "Site-directed mutagenesis of squalene–hopene cyclase: altered substrate specificity and product distribution". Chemistry & Biology. 7 (8): 643–649. doi:10.1016/S1074-5521(00)00003-X. ISSN 1074-5521. PMID 11048954.
- Rohmer, Michel; Anding, Claude; Ourisson, Guy (1980). "Non-specific Biosynthesis of Hopane Triterpenes by a Cell-Free System from Acetobacter pasteurianum". European Journal of Biochemistry. 112 (3): 541–547. doi:10.1111/j.1432-1033.1980.tb06117.x. ISSN 1432-1033. PMID 7460938.
- Perzl, Michael; Reipen, Ina G.; Schmitz, Susanne; Poralla, Karl; Sahm, Hermann; Sprenger, Georg A.; Kannenberg, Elmar L. (1998-07-31). "Cloning of conserved genes from Zymomonas mobilis and Bradyrhizobium japonicum that function in the biosynthesis of hopanoid lipids1EMBL accession number for the nucleotide sequence from Z. mobilis is AJ001401 and for the nucleotide sequence from B. japonicum, X86552.1". Biochimica et Biophysica Acta (BBA) - Lipids and Lipid Metabolism. 1393 (1): 108–118. doi:10.1016/S0005-2760(98)00064-2. ISSN 0005-2760. PMID 9714766.
- Bradley, Alexander S.; Pearson, Ann; Sáenz, James P.; Marx, Christopher J. (2010-10-01). "Adenosylhopane: The first intermediate in hopanoid side chain biosynthesis". Organic Geochemistry. 41 (10): 1075–1081. doi:10.1016/j.orggeochem.2010.07.003. ISSN 0146-6380.
- Liu, Wenjun; Sakr, Elias; Schaeffer, Philippe; Talbot, Helen M.; Donisi, Janina; Härtner, Thomas; Kannenberg, Elmar; Takano, Eriko; Rohmer, Michel (2014). "Ribosylhopane, a Novel Bacterial Hopanoid, as Precursor of C35 Bacteriohopanepolyols in Streptomyces coelicolor A3(2)". ChemBioChem. 15 (14): 2156–2161. doi:10.1002/cbic.201402261. ISSN 1439-7633. PMC 4245026. PMID 25155017.
- Schmerk, Crystal L.; Welander, Paula V.; Hamad, Mohamad A.; Bain, Katie L.; Bernards, Mark A.; Summons, Roger E.; Valvano, Miguel A. (2015). "Elucidation of the Burkholderia cenocepacia hopanoid biosynthesis pathway uncovers functions for conserved proteins in hopanoid-producing bacteria" (PDF). Environmental Microbiology. 17 (3): 735–750. doi:10.1111/1462-2920.12509. ISSN 1462-2920. PMID 24888970.
- Welander, Paula V.; Coleman, Maureen L.; Sessions, Alex L.; Summons, Roger E.; Newman, Dianne K. (2010-05-11). "Identification of a methylase required for 2-methylhopanoid production and implications for the interpretation of sedimentary hopanes". Proceedings of the National Academy of Sciences. 107 (19): 8537–8542. doi:10.1073/pnas.0912949107. ISSN 0027-8424. PMC 2889317. PMID 20421508.
- Welander, Paula V.; Summons, Roger E. (2012-08-07). "Discovery, taxonomic distribution, and phenotypic characterization of a gene required for 3-methylhopanoid production". Proceedings of the National Academy of Sciences. 109 (32): 12905–12910. doi:10.1073/pnas.1208255109. ISSN 0027-8424. PMC 3420191. PMID 22826256.
- Banta, Amy B.; Wei, Jeremy H.; Welander, Paula V. (2015-11-03). "A distinct pathway for tetrahymanol synthesis in bacteria". Proceedings of the National Academy of Sciences. 112 (44): 13478–13483. doi:10.1073/pnas.1511482112. ISSN 0027-8424. PMC 4640766. PMID 26483502.
- Ourisson, Guy; Albrecht, Pierre (September 1992). "Hopanoids. 1. Geohopanoids: the most abundant natural products on Earth?". Accounts of Chemical Research. 25 (9): 398–402. doi:10.1021/ar00021a003. ISSN 0001-4842.
- Summons, Roger E.; Lincoln, Sara A. (2012-03-30), "Biomarkers: Informative Molecules for Studies in Geobiology", Fundamentals of Geobiology, John Wiley & Sons, Ltd, pp. 269–296, doi:10.1002/9781118280874.ch15, ISBN 978-1-118-28087-4
- Knoll, Andrew H. (2003). Life on a young planet : the first three billion years of evolution on Earth. Princeton, N.J.: Princeton University Press. ISBN 0-691-00978-3. OCLC 50604948.
- Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E. (1999-08-13). "Archean Molecular Fossils and the Early Rise of Eukaryotes". Science. 285 (5430): 1033–1036. doi:10.1126/science.285.5430.1033. ISSN 0036-8075. PMID 10446042.
- Rashby, Sky E.; Sessions, Alex L.; Summons, Roger E.; Newman, Dianne K. (2007-09-18). "Biosynthesis of 2-methylbacteriohopanepolyols by an anoxygenic phototroph". Proceedings of the National Academy of Sciences. 104 (38): 15099–15104. doi:10.1073/pnas.0704912104. ISSN 0027-8424. PMC 1986619. PMID 17848515.
- Talbot, Helen M.; Summons, Roger E.; Jahnke, Linda L.; Cockell, Charles S.; Rohmer, Michel; Farrimond, Paul (2008-02-01). "Cyanobacterial bacteriohopanepolyol signatures from cultures and natural environmental settings". Organic Geochemistry. 39 (2): 232–263. doi:10.1016/j.orggeochem.2007.08.006. ISSN 0146-6380.
- French, Katherine L.; Hallmann, Christian; Hope, Janet M.; Schoon, Petra L.; Zumberge, J. Alex; Hoshino, Yosuke; Peters, Carl A.; George, Simon C.; Love, Gordon D.; Brocks, Jochen J.; Buick, Roger (2015-05-12). "Reappraisal of hydrocarbon biomarkers in Archean rocks". Proceedings of the National Academy of Sciences. 112 (19): 5915–5920. doi:10.1073/pnas.1419563112. ISSN 0027-8424. PMC 4434754. PMID 25918387.
- Brocks, Jochen J.; Love, Gordon D.; Summons, Roger E.; Knoll, Andrew H.; Logan, Graham A.; Bowden, Stephen A. (October 2005). "Biomarker evidence for green and purple sulphur bacteria in a stratified Palaeoproterozoic sea". Nature. 437 (7060): 866–870. doi:10.1038/nature04068. ISSN 1476-4687. PMID 16208367.
- Gold, David A.; Caron, Abigail; Fournier, Gregory P.; Summons, Roger E. (March 2017). "Paleoproterozoic sterol biosynthesis and the rise of oxygen". Nature. 543 (7645): 420–423. doi:10.1038/nature21412. ISSN 1476-4687. PMID 28264195.
- Ourisson, Guy; Albrecht, Pierre; Rohmer, Michel (1982-07-01). "Predictive microbial biochemistry — from molecular fossils to procaryotic membranes". Trends in Biochemical Sciences. 7 (7): 236–239. doi:10.1016/0968-0004(82)90028-7. ISSN 0968-0004.
- Fischer, W. W.; Pearson, A. (2007). "Hypotheses for the origin and early evolution of triterpenoid cyclases". Geobiology. 5 (1): 19–34. doi:10.1111/j.1472-4669.2007.00096.x. ISSN 1472-4669.
- Hammer, Stephan C.; Marjanovic, Antonija; Dominicus, Jörg M.; Nestl, Bettina M.; Hauer, Bernhard (February 2015). "Squalene hopene cyclases are protonases for stereoselective Brønsted acid catalysis". Nature Chemical Biology. 11 (2): 121–126. doi:10.1038/nchembio.1719. ISSN 1552-4469. PMID 25503928.
- , "Hopanoids producing bacteria and related biofertilizers, compositions, methods and systems", issued 2016-10-19
- Sáenz, James P.; Grosser, Daniel; Bradley, Alexander S.; Lagny, Thibaut J.; Lavrynenko, Oksana; Broda, Martyna; Simons, Kai (2015-09-22). "Hopanoids as functional analogues of cholesterol in bacterial membranes". Proceedings of the National Academy of Sciences. 112 (38): 11971–11976. doi:10.1073/pnas.1515607112. ISSN 0027-8424. PMC 4586864. PMID 26351677.