CASS4
Cas scaffolding protein family member 4 is a protein that in humans is encoded by the CASS4 gene.[5]


History and discovery
CASS4 (Crk associated substrate 4) is the fourth and last described member of the CAS protein family.[6] CASS4 was detected by Singh et al.[7] in 2008 following in silico screening of databases describing expressed sequence tags from an evolutionarily diverse group of organisms, using the CAS-related proteins (p130Cas, NEDD9/HEF1 and EFS) mRNAs as templates. Singh et al. subsequently cloned and characterized the CASS4 gene, originally assigning the name HEPL (HEF1-EFS-p130Cas-like) for similarity to the other three defined CAS genes. The official name was subsequently changed to CASS4 by the Human Genome Organization (HUGO) Gene Nomenclature Committee (HGNC).
Gene
The chromosomal location of the CASS4 gene is 20q13.31, with genomic coordinates of 20: 56411548-56459340 on the forward strand in GRChB38p2.[8] While its HGNC-approved symbol is CASS4, this gene has multiple synonyms, including "HEF-like protein", "HEF1-Efs-p130Cas-like", HEFL, HEPL and C20orf32 ("chromosome 20 open reading frame 32"). Official IDs assigned to this gene include 15878 (HGNC), 57091 (Entrez Gene) and ENSG00000087589 (Ensembl). In humans four transcript variants are known. The first and second each contain 7 exons and encode the same full-length protein isoform a (786 amino acids, considered the major isoform), the third one contains 6 exons and encodes a shorter isoform b (732 amino acids) and the fourth one contains 5 exons and encodes the shortest isoform c (349 amino acids). Cumulatively, the CASS4 transcripts are most highly expressed in spleen and lung among normal tissues, and are highly expressed in ovarian and leukemia cell lines.[7]
To date, little effort has been applied to the direct study of transcriptional regulation of CASS4. The SABiosciences’ DECODE database, based on the UCSC Bioinformatics Genome Browser,[9] proposes several transcriptional regulators for CASS4 based on its promoter region sequence: NF-κβ, p53, LCR-F1 (NFE2-L1, nuclear factor, erythroid 2-like1), MAX1, C/EBPα, CHOP-10 (C/EBP homologous protein 10), POU3F1 (POU domain, class 3, transcription factor 1, aka Oct-6), Areb6 (ZEB1, Zinc finger E-box binding homeobox 1). These are compatible with regulation relevant to lymphocytes and deregulation in cancer.
Protein family
In vertebrates, the CAS protein family contains four members: p130Cas/BCAR1, NEDD9/HEF1, EFS and CASS4. There are no paralogous genes for this family in acoelomates, pseudocoelomates, and nematodes, while a single ancestral member is found in Drosophila.[7] Evolutionary divergence of the CAS proteins family members is discussed by Singh et al. in detail.[7]
Structure
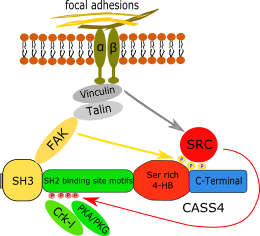
All CAS protein family members have common structural characteristics.[6] CAS proteins have an amino terminal SH3 domain enabling interaction with poly-proline motif-containing proteins such as FAK. Carboxy-terminal to this, they possess an unstructured domain containing multiple SH2 binding site motifs, which when tyrosine-phosphorylated allow interaction with SH2 domain containing proteins. Further to the carboxy-terminus, they have a four-helix bundle rich in serine residues, and a second highly conserved four-helix bundle that has been recognized as functionally and structurally similar to a focal adhesion targeting [FAT] domain.[8] For the better studied members of the CAS family (BCAR1 and NEDD9), all of these domains have been defined as crucial for recognition and binding by other proteins, reflecting the primary role of CAS family proteins as cell signaling cascades mediators.
Isoform “a” of human CASS4 is considered the predominant species, and at 786 amino acids is the longest one.[10] Amino acid sequence homology of this isoform of human CASS4 with other family members is 26% overall identity and 42% similarity.[7] Using a yeast two-hybrid approach, the CASS4 protein SH3 domain was shown to interact with the FAK C-terminus, despite the lowest overall similarity to other SH3 domains in the CAS group. In addition, human CASS4 has a limited number of candidate SH2-binding sites, estimated at 10, which is similar to EFS (estimated at 9) and in contrast to p130Cas/BCAR1 and NEDD9, which have 20 and 18 respectively. The CASS4 C-terminus has a short region of CAS family homology, but lacks obvious similarity at the level of primary amino acid sequence. It also lacks a YDYVHL sequence at the N-terminal end of the FAT-like carboxy-terminal domain, even though this motif is conserved among the other three CAS family proteins and is an important binding site for the Src SH2 domain.[11] Although this lack of sequence similarity may mean a reduced functionality of the CASS4 protein, molecular modeling analysis performed by Singh and colleagues[7] using p130CAS/BCAR1 structures as templates suggested an almost identical fold between CASS4 and p130CAS/BCAR1 within their SH3 domains, and substantial similarity within 432-591 residues of CASS4 and 449-610 residues of p130Cas/BCAR1 at the level of secondary and tertiary structures. Also, the similar periodicity of α-helices and β-sheets in both CASS4 and p130Cas/BCAR1 provides another confirmation for the idea of well-conserved structures within the family members.
Function
The exact function of CASS4 and its role in development and human pathologies have been subject to little investigation compared to other family members. The primary study exploring CASS4 function was the initial report by Singh et al.,[7] who showed the direct interaction between CASS4 and FAK, and CASS4 regulation of FAK activation, affecting cellular adhesion, migration and motility. Unusually, CASS4 depletion had a bimodal affect, causing some cells to have lower velocity and others to have higher velocity than control cells, suggesting a potential role in maintaining homeostasis. This work also suggested the function of CASS4 may be cell-type specific and dependent upon the presence or absence of expression of other CAS family members.[7] Direct binding has also been identified between CASS4 and CRKL,[12] an SH2- and SH3 domain-containing adaptor protein that has been also shown to interact with another CAS family member, p130Cas/BCAR1, in regulation of cellular motility and migration.[13] Because of the high degree of homology in interaction domains and some identified common partners, CASS4 is likely to share some functions with other CAS family members. These include association with FAK and Src family kinases at focal adhesions to transmit integrin-initiated signals to downstream effectors, which results in cytoskeleton reorganization and changes in motility and invasion.[14]
Disease association
Altered expression or modification of CASS4 has been proposed as relevant to several human pathologies, typically based on detection of changes in CASS4 in high throughput screening, although the role of CASS4 in the pathology of these conditions has not yet been studied directly. These findings are summarized in Table 1; some examples are provided below.
| Screen purpose | Observation |
|---|---|
| Alzheimer's disease | SNP rs7274581 T/C linked to risk. Odds ratio 0.72; p-value 0.011.[15] |
| SNP rs7274581 T/C linked to risk. Odds ratio 0.88; p-value 2.5*10−8.[16] | |
| SNP rs7274581 T/C linked to risk Odds ratio 0.8888; p-value 1.75 ×10−7.[17] | |
| SNP rs6024870, RegulomeDB[18] score 2b, which means that this SNP is likely to affect transcription factor binding.[19] | |
| SNP rs16979934 T/G linked to risk.
Odds ratio 0.5956; p-value 0.03.[20] | |
| Atopic asthma | Upregulated in eosinophils post antigen exposure.[21] |
| Cystic fibrosis | Possible correlation with severity of the lung manifestation of the disease (according to GWAS).[22] |
| Non-small cell lung cancer | Overexpression is associated with lymph node metastasis, high TNM stage and poor prognosis.[23] |
| Phosphoproteome of resting human platelets | Identified S305 phosphorylation by PKA/PKG in the unstructured region containing SH2-binding sites of CASS4.[24] |
| Platelet activation by oxidized phospholipids | CASS4 S249 phosphorylation in the unstructured region containing SH2-binding sites is induced by oxidized phospholipid KODA-PC in platelets.[25] |
Cancer
Many CAS family proteins have altered activity and functional roles in cancer progression and metastasis, with functional roles in influencing cellular adhesion, migration and drug resistance.[26][27] Changes in CASS4 may also be associated with human malignancies. CASS4 function was linked to non-small cell lung cancer (NSCLC) in a study by Miao et al. that correlated elevated CASS4 expression with lymph node metastasis and high TNM stage.[23] In addition, this study detected a significant difference in cytoplasmic accumulation of CASS4 protein between high (H1299 and BE1) and low (LTE and A549) metastatic potential lung cancer cell lines. These may suggest CASS4 as a possible prognostic marker in clinical management of NSCLC.
Alzheimer's disease
CASS4 and corresponding SNP - rs7274581 T/C has been identified in a large meta-analysis as a locus for lower susceptibility to Alzheimer's disease (AD).[16][28] However this SNP was not found predictive in a follow-up study.[17]
In a genome wide association screen (GWAS), CASS4 showed a significant correlation with clinical pathological features of AD such as neurofibrillary tangles and neuritic plaques.[15] Two additional CASS4 SNPs were reported to be associated with AD susceptibility: rs6024870,[19] and rs16979934 T/G.[20] Given the likely conserved CAS-family cytoskeletal function of CASS4, it has been speculated that it may have a role in axonal transport and influence the expression of the amyloid precursor protein (APP) and tau, which are pathologically affected in AD.[29] Several possible mechanisms for CASS4 action in AD have been proposed.[30]
Immunopathological conditions
An association of CASS4 with atopic asthma has been shown.[21] CASS4 has also been reported to be an eosinophil-associated gene, with expression in sputum cells increased more than 1.5-fold after whole lung allergen challenge. Moreover, the CASS4 mRNA was upregulated in cells collected by bronchoalveolar lavage after segmental broncho-provocation with an allergen. Reciprocally, the CASS4 mRNA was downregulated when this procedure was performed following administration of mepolizumab (a humanized monoclonal anti-IL-5 antibodies which reduces excessive eosinophilia). This suggests CASS4 activity may be associated with immune response in the context of atopic asthma development.
Cystic fibrosis
CASS4 has been reported to play a modifying role in cystic fibrosis severity, progression and comorbid conditions.[22] The CAS family member NEDD9 has also been shown to interact directly with AURKA (encoding Aurora-A kinase) to regulate cell cycle[31] and ciliary resorption;[32] it is possible that CASS4 may similarly interact with aurora-A kinase.
Thrombosis
CASS4 signaling may contribute to platelet activation and aggregation. A PKA/PKG phosphorylation site has been identified in CASS4 on residue S305 in the unstructured domain containing SH2-binding motifs; the functional significance of this phosphorylation is currently unknown.[24] Significantly increased phosphorylation on S249 of CASS4, also in the unstructured domain, after platelet stimulation with the oxidized phospholipid KODA-PC (9-keto-12-oxo-10-dodecenoic acid ester of 2-lyso-phosphocholine, a CD36 receptor agonist) versus thrombin treatment, which may implicate CASS4 mediated signaling in platelet hyperreactivity.[25]
Clinical significance
There are currently no therapeutic approaches targeting CASS4, and in the absence of a catalytic domain and no extracellular moieties, it may be challenging to generate such an agent. However, CASS4 may ultimately be relevant in clinical practice as a possible marker to assess prognosis and outcome in cases of NSCLC (and possibly other types of cancer). At present, its greatest clinical value is likely to be as a predictive variant for severity and onset of Alzheimer's disease and cystic fibrosis.
Notes
References
- GRCh38: Ensembl release 89: ENSG00000087589 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000074570 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: Cas scaffolding protein family member 4".
- Tikhmyanova N, Little JL, Golemis EA (April 2010). "CAS proteins in normal and pathological cell growth control". Cellular and Molecular Life Sciences. 67 (7): 1025–48. doi:10.1007/s00018-009-0213-1. PMC 2836406. PMID 19937461.
- Singh MK, Dadke D, Nicolas E, Serebriiskii IG, Apostolou S, Canutescu A, Egleston BL, Golemis EA (April 2008). "A novel Cas family member, HEPL, regulates FAK and cell spreading". Molecular Biology of the Cell. 19 (4): 1627–36. doi:10.1091/mbc.E07-09-0953. PMC 2291417. PMID 18256281.
- Tikhmyanova N, Golemis EA (2011). "NEDD9 and BCAR1 negatively regulate E-cadherin membrane localization, and promote E-cadherin degradation". PLOS ONE. 6 (7): e22102. doi:10.1371/journal.pone.0022102. PMC 3134485. PMID 21765937.
- "Search ChIP-qPCR Assay". sabiosciences. Archived from the original on 18 April 2015. Retrieved 17 April 2015.
- UniProt: Q9NQ75
- Tachibana K, Urano T, Fujita H, Ohashi Y, Kamiguchi K, Iwata S, Hirai H, Morimoto C (November 1997). "Tyrosine phosphorylation of Crk-associated substrates by focal adhesion kinase. A putative mechanism for the integrin-mediated tyrosine phosphorylation of Crk-associated substrates". The Journal of Biological Chemistry. 272 (46): 29083–90. doi:10.1074/jbc.272.46.29083. PMID 9360983.
- Brehme M, Hantschel O, Colinge J, Kaupe I, Planyavsky M, Köcher T, Mechtler K, Bennett KL, Superti-Furga G (May 2009). "Charting the molecular network of the drug target Bcr-Abl". Proceedings of the National Academy of Sciences of the United States of America. 106 (18): 7414–9. doi:10.1073/pnas.0900653106. PMC 2670881. PMID 19380743.
- Hsia DA, Mitra SK, Hauck CR, Streblow DN, Nelson JA, Ilic D, Huang S, Li E, Nemerow GR, Leng J, Spencer KS, Cheresh DA, Schlaepfer DD (March 2003). "Differential regulation of cell motility and invasion by FAK". The Journal of Cell Biology. 160 (5): 753–67. doi:10.1083/jcb.200212114. PMC 2173366. PMID 12615911.
- Tikhmyanova N, Tulin AV, Roegiers F, Golemis EA (2010). "Dcas supports cell polarization and cell-cell adhesion complexes in development". PLOS ONE. 5 (8): e12369. doi:10.1371/journal.pone.0012369. PMC 2927436. PMID 20808771.
- Beecham GW, Hamilton K, Naj AC, Martin ER, Huentelman M, Myers AJ, Corneveaux JJ, Hardy J, Vonsattel JP, Younkin SG, Bennett DA, De Jager PL, Larson EB, Crane PK, Kamboh MI, Kofler JK, Mash DC, Duque L, Gilbert JR, Gwirtsman H, Buxbaum JD, Kramer P, Dickson DW, Farrer LA, Frosch MP, Ghetti B, Haines JL, Hyman BT, Kukull WA, Mayeux RP, Pericak-Vance MA, Schneider JA, Trojanowski JQ, Reiman EM, Schellenberg GD, Montine TJ (September 2014). "Genome-wide association meta-analysis of neuropathologic features of Alzheimer's disease and related dementias". PLoS Genetics. 10 (9): e1004606. doi:10.1371/journal.pgen.1004606. PMC 4154667. PMID 25188341.
- Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, DeStafano AL, Bis JC, Beecham GW, Grenier-Boley B, Russo G, Thorton-Wells TA, Jones N, Smith AV, Chouraki V, Thomas C, Ikram MA, Zelenika D, Vardarajan BN, Kamatani Y, Lin CF, Gerrish A, Schmidt H, Kunkle B, Dunstan ML, Ruiz A, Bihoreau MT, Choi SH, Reitz C, Pasquier F, Cruchaga C, Craig D, Amin N, Berr C, Lopez OL, De Jager PL, Deramecourt V, Johnston JA, Evans D, Lovestone S, Letenneur L, Morón FJ, Rubinsztein DC, Eiriksdottir G, Sleegers K, Goate AM, Fiévet N, Huentelman MW, Gill M, Brown K, Kamboh MI, Keller L, Barberger-Gateau P, McGuiness B, Larson EB, Green R, Myers AJ, Dufouil C, Todd S, Wallon D, Love S, Rogaeva E, Gallacher J, St George-Hyslop P, Clarimon J, Lleo A, Bayer A, Tsuang DW, Yu L, Tsolaki M, Bossù P, Spalletta G, Proitsi P, Collinge J, Sorbi S, Sanchez-Garcia F, Fox NC, Hardy J, Deniz Naranjo MC, Bosco P, Clarke R, Brayne C, Galimberti D, Mancuso M, Matthews F, Moebus S, Mecocci P, Del Zompo M, Maier W, Hampel H, Pilotto A, Bullido M, Panza F, Caffarra P, Nacmias B, Gilbert JR, Mayhaus M, Lannefelt L, Hakonarson H, Pichler S, Carrasquillo MM, Ingelsson M, Beekly D, Alvarez V, Zou F, Valladares O, Younkin SG, Coto E, Hamilton-Nelson KL, Gu W, Razquin C, Pastor P, Mateo I, Owen MJ, Faber KM, Jonsson PV, Combarros O, O'Donovan MC, Cantwell LB, Soininen H, Blacker D, Mead S, Mosley TH, Bennett DA, Harris TB, Fratiglioni L, Holmes C, de Bruijn RF, Passmore P, Montine TJ, Bettens K, Rotter JI, Brice A, Morgan K, Foroud TM, Kukull WA, Hannequin D, Powell JF, Nalls MA, Ritchie K, Lunetta KL, Kauwe JS, Boerwinkle E, Riemenschneider M, Boada M, Hiltuenen M, Martin ER, Schmidt R, Rujescu D, Wang LS, Dartigues JF, Mayeux R, Tzourio C, Hofman A, Nöthen MM, Graff C, Psaty BM, Jones L, Haines JL, Holmans PA, Lathrop M, Pericak-Vance MA, Launer LJ, Farrer LA, van Duijn CM, Van Broeckhoven C, Moskvina V, Seshadri S, Williams J, Schellenberg GD, Amouyel P (December 2013). "Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease" (PDF). Nature Genetics. 45 (12): 1452–8. doi:10.1038/ng.2802. PMC 3896259. PMID 24162737.
- Ruiz A, Heilmann S, Becker T, Hernández I, Wagner H, Thelen M, Mauleón A, Rosende-Roca M, Bellenguez C, Bis JC, Harold D, Gerrish A, Sims R, Sotolongo-Grau O, Espinosa A, Alegret M, Arrieta JL, Lacour A, Leber M, Becker J, Lafuente A, Ruiz S, Vargas L, Rodríguez O, Ortega G, Dominguez MA, Mayeux R, Haines JL, Pericak-Vance MA, Farrer LA, Schellenberg GD, Chouraki V, Launer LJ, van Duijn C, Seshadri S, Antúnez C, Breteler MM, Serrano-Ríos M, Jessen F, Tárraga L, Nöthen MM, Maier W, Boada M, Ramírez A (2014). "Follow-up of loci from the International Genomics of Alzheimer's Disease Project identifies TRIP4 as a novel susceptibility gene". Translational Psychiatry. 4 (2): e358. doi:10.1038/tp.2014.2. PMC 3944635. PMID 24495969.
- "Search RegulomeDB". Regulomedb.org. Retrieved 30 January 2016.
- Rosenthal SL, Barmada MM, Wang X, Demirci FY, Kamboh MI (2014). "Connecting the dots: potential of data integration to identify regulatory SNPs in late-onset Alzheimer's disease GWAS findings". PLOS ONE. 9 (4): e95152. doi:10.1371/journal.pone.0095152. PMC 3990600. PMID 24743338.
- Wang X, Lopez OL, Sweet RA, Becker JT, DeKosky ST, Barmada MM, Demirci FY, Kamboh MI (2015). "Genetic determinants of disease progression in Alzheimer's disease". Journal of Alzheimer's Disease. 43 (2): 649–55. doi:10.3233/JAD-140729. PMC 4245313. PMID 25114068.
- Esnault S, Kelly EA, Schwantes EA, Liu LY, DeLain LP, Hauer JA, Bochkov YA, Denlinger LC, Malter JS, Mathur SK, Jarjour NN (2013). "Identification of genes expressed by human airway eosinophils after an in vivo allergen challenge". PLOS ONE. 8 (7): e67560. doi:10.1371/journal.pone.0067560. PMC 3699655. PMID 23844029.
- Wright FA, Strug LJ, Doshi VK, Commander CW, Blackman SM, Sun L, Berthiaume Y, Cutler D, Cojocaru A, Collaco JM, Corey M, Dorfman R, Goddard K, Green D, Kent JW, Lange EM, Lee S, Li W, Luo J, Mayhew GM, Naughton KM, Pace RG, Paré P, Rommens JM, Sandford A, Stonebraker JR, Sun W, Taylor C, Vanscoy LL, Zou F, Blangero J, Zielenski J, O'Neal WK, Drumm ML, Durie PR, Knowles MR, Cutting GR (June 2011). "Genome-wide association and linkage identify modifier loci of lung disease severity in cystic fibrosis at 11p13 and 20q13.2". Nature Genetics. 43 (6): 539–46. doi:10.1038/ng.838. PMC 3296486. PMID 21602797.
- Miao Y, Wang L, Liu Y, Li AL, Liu SL, Cao HY, Zhang XP, Jiang GY, Liu D, Wang EH (February 2013). "Overexpression and cytoplasmic accumulation of Hepl is associated with clinicopathological parameters and poor prognosis in non-small cell lung cancer". Tumour Biology. 34 (1): 107–14. doi:10.1007/s13277-012-0517-x. PMID 23001926.
- Zahedi RP, Lewandrowski U, Wiesner J, Wortelkamp S, Moebius J, Schütz C, Walter U, Gambaryan S, Sickmann A (February 2008). "Phosphoproteome of resting human platelets". Journal of Proteome Research. 7 (2): 526–34. doi:10.1021/pr0704130. PMID 18088087.
- Zimman A, Titz B, Komisopoulou E, Biswas S, Graeber TG, Podrez EA (2014). "Phosphoproteomic analysis of platelets activated by pro-thrombotic oxidized phospholipids and thrombin". PLOS ONE. 9 (1): e84488. doi:10.1371/journal.pone.0084488. PMC 3882224. PMID 24400094.
- Tornillo G, Defilippi P, Cabodi S (2014). "Cas proteins: dodgy scaffolding in breast cancer". Breast Cancer Research. 16 (5): 443. doi:10.1186/s13058-014-0443-5. PMC 4384296. PMID 25606587.
- Nikonova AS, Gaponova AV, Kudinov AE, Golemis EA (June 2014). "CAS proteins in health and disease: an update". IUBMB Life. 66 (6): 387–95. doi:10.1002/iub.1282. PMC 4111207. PMID 24962474.
- "IGAP Summary Statistics". Institut Pasteur de Lille.
- Karch CM, Goate AM (January 2015). "Alzheimer's disease risk genes and mechanisms of disease pathogenesis". Biological Psychiatry. 77 (1): 43–51. doi:10.1016/j.biopsych.2014.05.006. PMC 4234692. PMID 24951455.
- Beck TN, Nicolas E, Kopp MC, Golemis EA (2014). "Adaptors for disorders of the brain? The cancer signaling proteins NEDD9, CASS4, and PTK2B in Alzheimer's disease". Oncoscience. 1 (7): 486–503. doi:10.18632/oncoscience.64. PMC 4278314. PMID 25594051.
- Pugacheva EN, Golemis EA (October 2005). "The focal adhesion scaffolding protein HEF1 regulates activation of the Aurora-A and Nek2 kinases at the centrosome". Nature Cell Biology. 7 (10): 937–46. doi:10.1038/ncb1309. PMC 2652766. PMID 16184168.
- Pugacheva EN, Jablonski SA, Hartman TR, Henske EP, Golemis EA (June 2007). "HEF1-dependent Aurora A activation induces disassembly of the primary cilium". Cell. 129 (7): 1351–63. doi:10.1016/j.cell.2007.04.035. PMC 2504417. PMID 17604723.
External links
- Human CASS4 genome location and CASS4 gene details page in the UCSC Genome Browser.