Memory B cell
Memory B cells (MBCs) are a B cell sub-type that are formed within germinal centers following primary infection. Memory B cells can survive for decades and repeatedly generate an accelerated and robust antibody-mediated immune response in the case of re-infection (also known as a secondary immune response).[1][2]
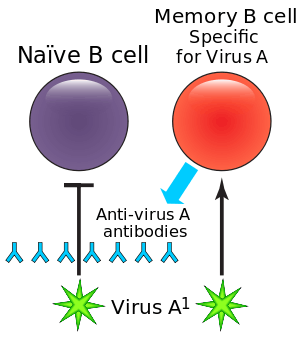
Primary response
In a T-cell dependent development pathway, naïve follicular B cells are activated by antigen presenting TFH cells during the initial infection, or primary immune response.[2] After activation, the B cells move into the secondary lymphoid organs (i.e. spleen and lymph nodes).[2] Within the secondary lymphoid organs, most of the B cells will enter B-cell follicles where a germinal center will form. Most B cells will eventually differentiate into plasma cells or memory B cells within the germinal center.[2][3]
Once inside the germinal center, the B cells undergo proliferation, followed by mutation of the genetic coding region of their surface receptors, a process known as somatic hypermutation.[2] The mutations will either increase or decrease the affinity of the surface receptor for a particular antigen, a progression called affinity maturation. After acquiring these mutations, the receptors on the surface of the B cells (B cell receptors) are tested within the germinal center for their affinity to the current antigen.[4] B cell clones with mutations that have increased the affinity of their surface receptors receive survival signals via interactions with their cognate TFH cells.[1][2][5] The B cells that do not have high enough affinity to receive these survival signals, as well as B cells that are potentially auto-reactive, will be selected against and die through apoptosis.[3] In addition to somatic hypermutation, many B cells will also undergo class switching before differentiation, which allows them to secrete different types of antibodies in future immune responses.[2]
Many B cells will differentiate into the plasma cells, also called effector B cells, which produce a first wave of protective antibodies and help clear infection.[3][1] A fraction of the B cells differentiate into memory B cells that survive long-term in the body.[6] The process of differentiation into memory B cells within the germinal center is not yet fully understood.[2] Some researchers hypothesize that differentiation into memory B cells occurs randomly.[3][7] Other hypotheses propose that the transcription factor NF-κB and the cytokine IL-24 are involved in the process of differentiation into memory B cells.[8][2] An additional hypothesis states that the B cells with relatively lower affinity for antigen will become memory B cells, in contrast to B cells with relatively higher affinity that will become plasma cells.[3][8]
After differentiation, memory B cells relocate to the periphery of the body where they will be more likely to encounter antigen in the event of a future exposure.[3][1][2] Many of the circulating B cells become concentrated in areas of the body that have a high likelihood of coming into contact with antigen, such as the Peyer's patch.[2]
Secondary response and memory
The memory B cells produced during the primary immune response are specific to the antigen involved during the first exposure. In a secondary response, the memory B cells specific to the antigen or similar antigens will respond.[2] When memory B cells reencounter their specific antigen, they proliferate and differentiate into plasma cells, which then respond to and clear the antigen.[2] The memory B cells that do not differentiate into plasma cells at this point can reenter the germinal centers to undergo further class switching or somatic hypermutation for further affinity maturation.[2] Differentiation of memory B cells into plasma cells is far faster than differentiation by naïve B cells, which allows memory B cells to produce a more efficient secondary immune response.[7] The efficiency and accumulation of the memory B cell response is the foundation for vaccines and booster shots.[7][2]
Lifespan
Memory B cells can survive for decades, which gives them the capacity to respond to multiple exposures to the same antigen.[2] The long-lasting survival is hypothesized to be a result of certain anti-apoptosis genes that are more highly expressed in memory B cells than other subsets of B cells.[3] Additionally, the memory B cell does not need to have continual interaction with the antigen nor with T cells in order to survive long-term.[7]
Markers
Memory B cells are typically distinguished by the cell surface marker CD27, although some subsets do not express CD27. Memory B cells that lack CD27 are generally associated with exhausted B cells or certain autoimmune conditions such as HIV, lupus, or rheumatoid arthritis.[1][2]
Because B cells have typically undergone class switching, they can express a range of immunoglobulin molecules. Some specific attributes of particular immunoglobulin molecules are described below:
- IgM: Memory B cells that express IgM can be found concentrated in the tonsils, Peyer's patch, and lymph nodes.[2] This subset of memory B cells is more likely to proliferate and reenter the germinal center during a secondary immune response.[7]
- IgG: Memory B cells that express IgG typically differentiate into plasma cells.[7]
- IgE: Memory B cells that express IgE are very rare in healthy individuals. This may occur because B cells that express IgE more frequently differentiate into plasma cells rather than memory B cells [7]
- IgD only: Memory B cells that express IgD are very rare. B cells with only IgD are found concentrated in the tonsils.[2]
The receptor CCR6 is generally a marker of B cells that will eventually differentiate into MBCs. This receptor detects chemokines, which are chemical messengers that allow the B cell to move within the body. Memory B cells may have this receptor to allow them to move out of the germinal center and into the tissues where they have a higher probability of encountering antigen.[3]
Subsets
Germinal center independent memory B cells
This subset of cells differentiates from activated B cells into memory B cells before entering the germinal center. B cells that have a high level of interaction with TFH within the B cell follicle have a higher propensity of entering the germinal center. The B cells that develop into memory B cells independently from germinal centers likely experience CD40 and cytokine signaling from T cells.[7] Class switching can still occur prior to interaction with the germinal center, while somatic hypermutation only occurs after interaction with the germinal center.[7] The lack of somatic hypermutation is hypothesized to be beneficial; a lower level of affinity maturation means that these memory B cells are less specialized to a specific antigen and may be able to recognize a wider range of antigens.[8][9][7]
T-independent memory B cells
T-independent memory B cells are a subset called B1 cells. These cells generally reside in the peritoneal cavity. When reintroduced to antigen, some of these B1 cells can differentiate into memory B cells without interacting with a T cell.[7] These B cells produce IgM antibodies to help clear infection.[10]
T-bet memory B cells
T-bet B cells are a subset that have been found to express the transcription factor T-bet. T-bet is associated with class switching. T-bet B cells are also thought to be important in immune responses against intracellular bacterial and viral infections.[11]
See also
References
- Weisel, Florian; Shlomchik, Mark (2017-04-26). "Memory B Cells of Mice and Humans". Annual Review of Immunology. 35 (1): 255–284. doi:10.1146/annurev-immunol-041015-055531. ISSN 0732-0582. PMID 28142324.
- Seifert, M; Küppers, R (2016-08-08). "Human memory B cells". Leukemia. 30 (12): 2283–2292. doi:10.1038/leu.2016.226. ISSN 0887-6924. PMID 27499139.
- Suan, Dan; Sundling, Christopher; Brink, Robert (2017-04-01). "Plasma cell and memory B cell differentiation from the germinal center". Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology. 45: 97–102. doi:10.1016/j.coi.2017.03.006. ISSN 0952-7915. PMID 28319733.
- Allman, David; Wilmore, Joel R.; Gaudette, Brian T. (March 2019). "The continuing story of T‐cell independent antibodies". Immunological Reviews. 288 (1): 128–135. doi:10.1111/imr.12754. ISSN 0105-2896. PMC 6653682. PMID 30874357.
- Victora, Gabriel D.; Nussenzweig, Michel C. (2012-03-26). "Germinal Centers". Annual Review of Immunology. 30 (1): 429–457. doi:10.1146/annurev-immunol-020711-075032. ISSN 0732-0582. PMID 22224772.
- Gatto, Dominique; Brink, Robert (2010-11-01). "The germinal center reaction". Journal of Allergy and Clinical Immunology. 126 (5): 898–907. doi:10.1016/j.jaci.2010.09.007. ISSN 0091-6749. PMID 21050940.
- Kurosaki, Tomohiro; Kometani, Kohei; Ise, Wataru (March 2015). "Memory B cells". Nature Reviews Immunology. 15 (3): 149–159. doi:10.1038/nri3802. ISSN 1474-1733. PMID 25677494.
- Shinnakasu, Ryo; Kurosaki, Tomohiro (2017-04-01). "Regulation of memory B and plasma cell differentiation". Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology. 45: 126–131. doi:10.1016/j.coi.2017.03.003. ISSN 0952-7915. PMID 28359033.
- Pupovac, Aleta; Good-Jacobson, Kim L (2017-04-01). "An antigen to remember: regulation of B cell memory in health and disease". Current Opinion in Immunology. Lymphocyte development and activation * Tumour immunology. 45: 89–96. doi:10.1016/j.coi.2017.03.004. ISSN 0952-7915. PMC 7126224. PMID 28319732.
- Montecino-Rodriguez, Encarnacion; Dorshkind, Kenneth (2012-01-27). "B-1 B Cell Development in the Fetus and Adult". Immunity. 36 (1): 13–21. doi:10.1016/j.immuni.2011.11.017. ISSN 1074-7613. PMC 3269035. PMID 22284417.
- Knox, James J.; Myles, Arpita; Cancro, Michael P. (March 2019). "T‐bet + memory B cells: Generation, function, and fate". Immunological Reviews. 288 (1): 149–160. doi:10.1111/imr.12736. ISSN 0105-2896. PMC 6626622. PMID 30874358.