Neurexin
Neurexins (NRXN) are a family of presynaptic cell adhesion proteins that have roles in connecting neurons at the synapse.[1] They are located mostly on the presynaptic membrane and contain a single transmembrane domain. The extracellular domain interacts with proteins in the synaptic cleft, most notably neuroligin, while the intracellular cytoplasmic portion interacts with proteins associated with exocytosis.[2] Neurexin and neuroligin "shake hands," resulting in the connection between the two neurons and the production of a synapse.[3] Neurexins mediate signaling across the synapse, and influence the properties of neural networks by synapse specificity. Neurexins were discovered as receptors for α-latrotoxin, a vertebrate-specific toxin in black widow spider venom that binds to presynaptic receptors and induces massive neurotransmitter release.[4] In humans, alterations in genes encoding neurexins are implicated in autism and other cognitive diseases, such as Tourette syndrome and schizophrenia.[4]
| Neurexin family | |
|---|---|
| Identifiers | |
| Symbol | NRXN1_fam |
| InterPro | IPR037440 |
| Membranome | 15 |
| neurexin 1 | |
|---|---|
 3D ribbon diagram of alpha-neurexin 1 | |
| Identifiers | |
| Symbol | NRXN1 |
| NCBI gene | 9378 |
| HGNC | 8008 |
| OMIM | 600565 |
| RefSeq | NM_001135659.1 |
| UniProt | Q9ULB1 |
| Other data | |
| Locus | Chr. 2 p16.3 |
| neurexin 2 | |
|---|---|
| Identifiers | |
| Symbol | NRXN2 |
| NCBI gene | 9379 |
| HGNC | 8009 |
| OMIM | 600566 |
| RefSeq | NM_015080 |
| UniProt | P58401 |
| Other data | |
| Locus | Chr. 2 q13.1 |
| neurexin 3 | |
|---|---|
| Identifiers | |
| Symbol | NRXN3 |
| NCBI gene | 9369 |
| HGNC | 8010 |
| OMIM | 600567 |
| RefSeq | NM_001105250 |
| UniProt | Q9HDB5 |
| Other data | |
| Locus | Chr. 14 q31 |
| neurexin | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Organism | |||||||
| Symbol | Nrx-IV | ||||||
| Entrez | 39387 | ||||||
| RefSeq (mRNA) | NM_168491.3 | ||||||
| RefSeq (Prot) | NP_524034.2 | ||||||
| UniProt | Q94887 | ||||||
| Other data | |||||||
| Chromosome | 3L: 12.14 - 12.15 Mb | ||||||
| |||||||
| neurexin | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Organism | |||||||
| Symbol | Nrxn1 | ||||||
| Entrez | 18189 | ||||||
| RefSeq (mRNA) | NM_177284.2 | ||||||
| RefSeq (Prot) | NP_064648.3 | ||||||
| UniProt | Q9CS84 | ||||||
| Other data | |||||||
| Chromosome | 17: 90.03 - 91.09 Mb | ||||||
| |||||||
Structure
In mammals, neurexin is encoded by three different genes (NRXN1, NRXN2, and NRXN3) each controlled by two different promoters, an upstream alpha (α) and a downstream beta (β), resulting in alpha-neurexins 1-3 (α-neurexins 1-3) and beta-neurexins 1-3 (β-neurexins 1-3).[5] In addition, there are alternative splicing at 5 sites in α-neurexin and 2 in β-neurexin; more than 2000 splice variants are possible, suggesting its role in determining synapse specificity.[6]
The encoded proteins are structurally similar to laminin, slit, and agrin, other proteins involved in axon guidance and synaptogenesis.[6] α-Neurexins and β-neurexins have identical intracellular domains but different extracellular domains. The extracellular domain of α-neurexin is composed of three neurexin repeats which each contain LNS (laminin, neurexin, sex-hormone binding globulin) – EGF (epidermal growth factor) – LNS domains. N1α binds to a variety of ligands including neuroligins and GABA receptors,[2] though neurons of every receptor type express neurexins. β-Neurexins are shorter versions of α-neurexins, containing only one LNS domain.[7] β-Neurexins (located presynaptically) act as receptors for neuroligin (located postsynaptically). Additionally, β-Neurexin has also been found to play a role in angiogenesis.[8]
The C terminus of the short intracellular section of both types of neurexins binds to synaptotagmin and to the PDZ (postsynaptic density (PSD)-95/discs large/zona-occludens-1) domains of CASK and Mint. These interactions form connections between intracellular synaptic vesicles and fusion proteins.[9] Thus neurexins play an important role in assembling presynaptic and postsynaptic machinery.
Trans-synapse, the extracellular LNS domains have a functional region, the hyper-variable surface, formed by loops carrying 3 splice inserts.[2] This region surrounds a coordinated Ca2+ ion and is the site of neuroligin binding,[9] resulting in a neurexin-neuroligin Ca2+-dependent complex at the junction of chemical synapses.[10]
Expression and function
Neurexins are diffusely distributed in neurons and become concentrated at presynaptic terminals as neurons mature. There exists a trans-synaptic dialog between neurexin and neuroligin, meaning neuroligin can induce the expression of neurexin and vice versa.[11] This bi-directional trigger aids in the formation of synapses and is a key component to modifying the neuronal network. Over-expression of either of these proteins causes an increase in synapse forming sites, thus providing evidence that neurexin plays a functional role in synaptogenesis.[7] Conversely, the blocking of β-neurexin interactions reduces the number of excitatory and inhibitory synapses. It is not clear how exactly neurexin promotes the formation of synapses. One possibility is that actin is polymerized on the tail end of β-neurexin, which traps and stabilizes accumulating synaptic vesicles. This forms a forward feeding cycle, where small clusters of β-neurexins recruit more β-neurexins and scaffolding proteins to form a large synaptic adhesive contact.[7]
Neurexin-neuroligin binding
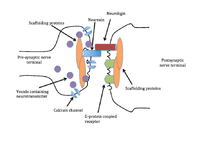
The different combinations of neurexin to neuroligin, and alternative splicing of neuroligin and neurexin genes, control binding between neuroligins and neurexins, adding to synapse specificity.[7] Neurexins alone are capable of recruiting neuroligins in postsynaptic cells to a dendritic surface, resulting in clustered neurotransmitter receptors and other postsynaptic proteins and machinery. Their neuroligin partners can induce presynaptic terminals by recruiting neurexins. Synapse formation can therefore be triggered in either direction by these proteins.[9] Neuroligins and neurexins can also regulate formation of glutamatergic (excitatory) synapses, and GABAergic (inhibitory) contacts using a neuroligin link. Regulating these contacts suggests neurexin-neuroligin binding could balance synaptic input,[6] or maintain an optimal ratio of excitatory to inhibitory contacts.
Additional interacting partners
Neurexins not only bind to neuroligin. Additional binding partners of neurexin are dystroglycan and neuroexophilins.[9] Dystroglycan is Ca2+-dependant and binds preferentially to α-neurexins on LNS domains that lack splice inserts. In mice, a deletion of dystroglycan causes long-term potentiation impairment and developmental abnormalities similar to muscular dystrophy; however baseline synaptic transmission is normal. Neuroexophilins are Ca2+-independent and bind exclusively to α-neurexins on the second LNS domain. The increased startle responses and impaired motor coordination of neuroexophilin knockout mice indicates that neuroexophilins have a functional role in certain circuits.[9] The significance of the relationship between neurexin and dystroglycan or neuroexophilins is still unclear.
Species distribution
Members of the neurexin family are found across all animals, including basal metazoans such as porifera (sponges), cnidaria (jellyfish) and ctenophora (comb jellies). Porifera lack synapses so its role in these organisms is unclear.
Homologues of α-neurexin have also been found in several invertebrate species including Drosophila, Caenorhabditis elegans, honeybees and Aplysia.[11] In Drosophila melanogaster, NRXN genes (only one α-neurexin) are critical in the assembly of glutamatergic neuromuscular junctions but are much simpler.[5] Their functional roles in insects are likely similar to those in vertebrates.[12]
Role in synaptic maturation
Neurexin and neuroligin have been found to be active in synapse maturation and adaptation of synaptic strength. Studies in knockout mice show that the trans-synaptic binding team does not increase the number of synaptic sites, but rather increases the strength of the existing synapses.[11] Deletion of the neurexin genes in the mice significantly impaired synaptic function, but did not alter synaptic structure. This is attributed to the impairment of specific voltage gated ion channels. While neuroligin and neurexin are not required for synaptic formation, they are essential components for proper function.[11]
Clinical importance and applications
Recent studies link mutations in genes encoding neurexin and neuroligin to a spectrum of cognitive disorders, such as autism spectrum disorders (ASDs), schizophrenia, and mental retardation.[4] Cognitive diseases remain difficult to understand, as they are characterized by subtle changes in a subgroup of synapses in a circuit rather than impairment of all systems in all circuits. Depending on the circuit, these subtle synapse changes may produce different neurological symptoms, leading to classification of different diseases. Counterarguments to the relationship between cognitive disorders and these mutations exist, prompting further investigation into the underlying mechanisms producing these cognitive disorders.
Autism
Autism is a neurodevelopmental disorder characterized by qualitative deficits in social behavior and communication, often including restricted, repetitive patterns of behavior.[13] It includes a subset of three disorders: childhood disintegrative disorder (CDD), Asperger syndrome (AS), and pervasive developmental disorder – not otherwise specified (PDD-NOS). A small percentage of ASD patients present with single mutations in genes encoding neuroligin-neurexin cell adhesion molecules. Neurexin is crucial to synaptic function and connectivity, as highlighted in wide spectrum of neurodevelopmental phenotypes in individuals with neurexin deletions. This provides strong evidence that neurexin deletions result in increased risk of ASDs, and indicate synapse dysfunction as the possible site of autism origin.[14] Dr. Steven Clapcote et al.'s α-neurexin II (Nrxn2α) KO mice experiments demonstrate a causal role for the loss of Nrxn2α in the genesis of autism-related behaviors in mice.[15]
Schizophrenia
Schizophrenia is a debilitating neuropsychiatric illness with multiple genes and environmental exposures involved in its genesis.[16] Further research indicates that deletion of the NRXN1 gene increases the risk of schizophrenia.[17] Genomic duplications and deletions on a micro-level – known as copy number variants (CNVs) – often underlie neurodevelopmental syndromes. Genomic-wide scans suggest that individuals with schizophrenia have rare structural variants that deleted or duplicated one or more genes.[16] As these studies only indicate an increased risk, further research is necessary to elucidate the underlying mechanisms of the genesis of cognitive diseases.[18]
Intellectual disability and Tourette syndrome
Similar to schizophrenia, studies have shown that intellectual disability and Tourette syndrome are also associated with NRXN1 deletions.[4][16] A recent study shows that NRXN genes 1-3 are essential for survival and play a pivotal and overlapping role with each other in neurodevelopment. These genes have been directly disrupted in Tourette syndrome by independent genomic rearrangements.[19] Another study suggests that NLGN4 mutations can be associated with a wide spectrum of neuropsychiatric conditions and that carriers may be affected with milder symptoms.[20]
See also
References
- Li X, Zhang J, Cao Z, Wu J, Shi Y (September 2006). "Solution structure of GOPC PDZ domain and its interaction with the C-terminal motif of neuroligin". Protein Sci. 15 (9): 2149–58. doi:10.1110/ps.062087506. PMC 2242614. PMID 16882988.
- Chen F, Venugopal V, Murray B, Rudenko G (June 2011). "The structure of neurexin 1α reveals features promoting a role as synaptic organizer". Structure. 19 (6): 779–89. doi:10.1016/j.str.2011.03.012. PMC 3134934. PMID 21620716.
- Scheiffele P, Fan J, Choih J, Fetter R, Serafini T (June 2000). "Neuroligin expressed in nonneuronal cells triggers presynaptic development in contacting axons". Cell. 101 (6): 657–69. doi:10.1016/S0092-8674(00)80877-6. PMID 10892652.
- Südhof TC (October 2008). "Neuroligins and neurexins link synaptic function to cognitive disease". Nature. 455 (7215): 903–11. Bibcode:2008Natur.455..903S. doi:10.1038/nature07456. PMC 2673233. PMID 18923512.
- Baudouin S, Scheiffele P (May 2010). "SnapShot: Neuroligin-neurexin complexes". Cell. 141 (5): 908–908.e1. doi:10.1016/j.cell.2010.05.024. PMID 20510934.
- Binder MD (2009). Encyclopedia of Neuroscience: Neurexins. Springer Berlin Heidelberg. p. 2607. ISBN 978-3-540-29678-2.
- Dean C, Dresbach T (January 2006). "Neuroligins and neurexins: linking cell adhesion, synapse formation and cognitive function". Trends Neurosci. 29 (1): 21–9. doi:10.1016/j.tins.2005.11.003. PMID 16337696.
- Bottos A, Destro E, Rissone A, Graziano S, Cordara G, Assenzio B, Cera MR, Mascia L, Bussolino F, Arese M (December 2009). "The synaptic proteins neurexins and neuroligins are widely expressed in the vascular system and contribute to its functions". Proc. Natl. Acad. Sci. U.S.A. 106 (49): 20782–7. Bibcode:2009PNAS..10620782B. doi:10.1073/pnas.0809510106. PMC 2791601. PMID 19926856.
- Craig AM, Kang Y (February 2007). "Neurexin-neuroligin signaling in synapse development". Curr. Opin. Neurobiol. 17 (1): 43–52. doi:10.1016/j.conb.2007.01.011. PMC 2820508. PMID 17275284.
- Reissner C, Klose M, Fairless R, Missler M (September 2008). "Mutational analysis of the neurexin/neuroligin complex reveals essential and regulatory components". Proc. Natl. Acad. Sci. U.S.A. 105 (39): 15124–9. Bibcode:2008PNAS..10515124R. doi:10.1073/pnas.0801639105. PMC 2551626. PMID 18812509.
- Knight D, Xie W, Boulianne GL (December 2011). "Neurexins and neuroligins: recent insights from invertebrates". Mol. Neurobiol. 44 (3): 426–40. doi:10.1007/s12035-011-8213-1. PMC 3229692. PMID 22037798.
- Biswas S, Russell RJ, Jackson CJ, Vidovic M, Ganeshina O, Oakeshott JG, Claudianos C (2008). "Bridging the synaptic gap: neuroligins and neurexin I in Apis mellifera". PLOS ONE. 3 (10): e3542. Bibcode:2008PLoSO...3.3542B. doi:10.1371/journal.pone.0003542. PMC 2570956. PMID 18974885.
- Lord C, Cook EH, Leventhal BL, Amaral DG (November 2000). "Autism spectrum disorders". Neuron. 28 (2): 355–63. doi:10.1016/S0896-6273(00)00115-X. PMID 11144346.
- Pizzarelli R, Cherubini E (2011). "Alterations of GABAergic signaling in autism spectrum disorders". Neural Plast. 2011: 297153. doi:10.1155/2011/297153. PMC 3134996. PMID 21766041.
- J Dachtler; J Glaspe (2014). "Deletion of α-neurexin II results in autism-related behaviors in mice". Translational Psychiatry. 4 (11): e484. doi:10.1038/tp.2014.123. PMC 4259993. PMID 25423136.
- Walsh T, McClellan JM, McCarthy SE, Addington AM, Pierce SB, Cooper GM, Nord AS, Kusenda M, Malhotra D, Bhandari A, Stray SM, Rippey CF, Roccanova P, Makarov V, Lakshmi B, Findling RL, Sikich L, Stromberg T, Merriman B, Gogtay N, Butler P, Eckstrand K, Noory L, Gochman P, Long R, Chen Z, Davis S, Baker C, Eichler EE, Meltzer PS, Nelson SF, Singleton AB, Lee MK, Rapoport JL, King MC, Sebat J (April 2008). "Rare structural variants disrupt multiple genes in neurodevelopmental pathways in schizophrenia". Science. 320 (5875): 539–43. Bibcode:2008Sci...320..539W. doi:10.1126/science.1155174. PMID 18369103.
- Kirov G, Rujescu D, Ingason A, Collier DA, O'Donovan MC, Owen MJ (September 2009). "Neurexin 1 (NRXN1) deletions in schizophrenia". Schizophr Bull. 35 (5): 851–4. doi:10.1093/schbul/sbp079. PMC 2728827. PMID 19675094.
- Kirov G, Gumus D, Chen W, Norton N, Georgieva L, Sari M, O'Donovan MC, Erdogan F, Owen MJ, Ropers HH, Ullmann R (February 2008). "Comparative genome hybridization suggests a role for NRXN1 and APBA2 in schizophrenia". Hum. Mol. Genet. 17 (3): 458–65. doi:10.1093/hmg/ddm323. PMID 17989066.
- Boschero AC, Negreiros de Paiva CE (1977). "Transplantation of islets of Langerhans in diabetic rats". Acta Physiol Lat Am. 27 (1): 1–6. PMID 356520.
- Kuttig P (September 1970). "[Social service for paraplegics and childhood paralytics]". Dtsch Schwesternztg (in German). 23 (9): 450–2. PMID 5202006.