Reeler
A reeler is a mouse mutant, so named because of its characteristic "reeling" gait. This is caused by the profound underdevelopment of the mouse's cerebellum, a segment of the brain responsible for locomotion. The mutation is autosomal and recessive, and prevents the typical cerebellar folia from forming.
Cortical neurons are generated normally but are abnormally placed, resulting in disorganization of cortical laminar layers in the central nervous system. The reason is the lack of Reelin, an extracellular matrix glycoprotein, which, during the corticogenesis, is secreted mainly by the Cajal-Retzius cells. In the reeler neocortex, cortical plate neurons are aligned in a practically inverted fashion (‘‘outside-in’’). In the ventricular zone of the cortex fewer neurons have been found to have radial glial processes.[1] In the dentate gyrus of hippocampus, no characteristic radial glial scaffold is formed and no compact granule cell layer is established.[2] Therefore, the reeler mouse presents a good model in which to investigate the mechanisms of establishment of the precise neuronal network during development.
Types of reelers
There are two types of the reeler mutation:
- Albany2 mutation (Reln(rl-Alb2)[3]
- Orleans mutation (Reln-rl-orl, or rl-orl),[4] in which reelin lacks a C-terminal region and a part of the eighth reelin repeat. This hampers the secretion of the protein from the cell.
In order to unravel the reelin signaling chain, attempts are made to cut the signal downstream of reelin, leaving reelin expression intact but creating the reeler phenotype, sometimes a partial phenotype, thus confirming the role of downstream molecules. The examples include:
- Double knockout of VLDLR and ApoER2 receptors;[5]
- Double knockout of Src and Fyn kinases.[6]
- Cre-loxP recombination mice model that lacks Crk and CrkL in most neurons.[7] Was used to show that Crk/CrkL lie downstream of DAB1] in the reelin signaling pathway.
- Scrambler mouse
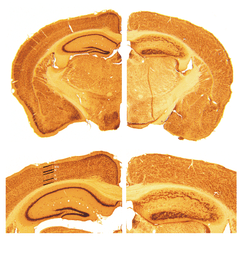
Key pathological findings in the Reeler brain structure
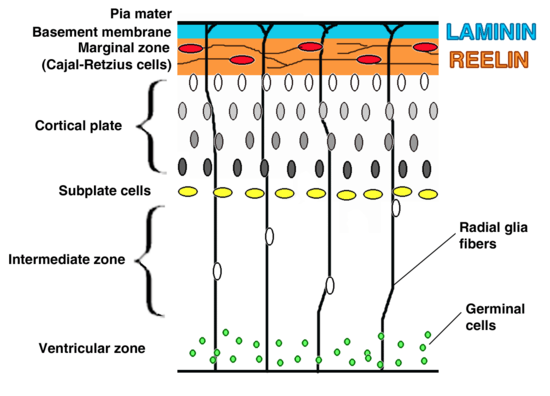
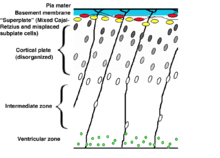
- Inversion of cortical layers.
- Subplate cells become abnormally located in the subpial zone above the cortical plate. This hampers their function in establishing the transient circuits between the incoming thalamic axons and layer IV cells of the cortical plate. Thalamic axons have to grow past the cortical plate to reach the mispositioned subplate cells in the so-called superplate and then turn back down to contact their appropriate targets.[8][9][10] This creates a curious "looping" thalamocortical connection seen in the adult reeler brain.
- Dispersion of neurons within cortical layers.
- Decreased cerebellar size.
- Failure of preplate to split
- Failure to establish a distinct granule cell layer in the dentate gyrus. Normal dentate gyrus demonstrates a clear segregation of granule cells and hilar mossy cells, which are identified, respectively, by their expression of calbindin and calretinin.[11] In the reeler DG, the two cell types intermingle.[12]
- Impaired dendrite outgrowth.[13]
- In one study, reeler mice were shown to have attenuated methamphetamine-induced hyperlocomotion, which was also reduced by a targeted disruption of reelin activity in wildtype mice. Reeler mice in the same study demonstrated a decrease in D1 and D2 receptor-mediated dopaminergic function together with reduced numbers of D1\D2 receptors. [14]
Heterozygous Reeler Mouse
Heterozygous reeler mice, also known as HRM, while lacking the apparent phenotype seen in the homozygous reeler, also show some brain abnormalities due to the reelin deficit.
Heterozygous (rl/+) mice express reelin at 50% of wild-type levels and have grossly normal brains but exhibit a progressive loss during aging of a neuronal target of reelin action, Purkinje cells.[15]
The mice have reduced density of parvalbumin-containing interneurons in circumscribed regions of striatum, according to one study.[16]
Studies reveal a 16% deficit in the number of Purkinje cells in 3-month-old (+/rl) and a 24% one in 16-month-old animals: surprisingly this deficit is only present in the (+/rl) males, while the females are spared.
History of research
First mention of reeler mouse mutation dates back to 1951.[17] In the later years, histopathological studies revealed that the reeler cerebellum is dramatically decreased in size and the normal laminar organization found in several brain regions is disrupted (Hamburgh, 1960). In 1995, the RELN gene and reelin protein were discovered at chromosome 7q22 by Tom Curran and colleagues.[18]
See also
- Shaking rat Kawasaki has a reduced reelin expression due to missplicing of the reelin gene.
- Sticky mouse
References
- Hartfuss E, Forster E, Bock HH, Hack MA, Leprince P, Luque JM, Herz J, Frotscher M, Gotz M (2003). "Reelin signaling directly affects radial glia morphology and biochemical maturation". Development. 130 (19): 4597–609. doi:10.1242/dev.00654. PMID 12925587.
- Weiss KH, Johanssen C, Tielsch A, Herz J, Deller T, Frotscher M, Förster E (2003). "Malformation of the radial glial scaffold in the dentate gyrus of reeler mice, scrambler mice, and ApoER2/VLDLR-deficient mice". J. Comp. Neurol. 460: 56–65. doi:10.1002/cne.10644. PMID 12687696.
- Royaux I, Bernier B, Montgomery JC, Flaherty L, Goffinet AM (1997). "Reln(rl-Alb2), an allele of reeler isolated from a chlorambucil screen, is due to an IAP insertion with exon skipping". Genomics. 42 (3): 479–82. doi:10.1006/geno.1997.4772. PMID 9205121.
- Lalonde R, Hayzoun K, Derer M, Mariani J, Strazielle C (2004). "Neurobehavioral evaluation of Reln-rl-orl mutant mice and correlations with cytochrome oxidase activity". Neurosci. Res. 49 (3): 297–305. doi:10.1016/j.neures.2004.03.012. PMID 15196778.
- Trommsdorff M, Gotthardt M, Hiesberger T, Shelton J, Stockinger W, Nimpf J, Hammer RE, Richardson JA, Herz J (June 1999). "Reeler/Disabled-like disruption of neuronal migration in knockout mice lacking the VLDL receptor and ApoE receptor 2". Cell. 97 (6): 689–701. doi:10.1016/S0092-8674(00)80782-5. PMID 10380922.
- Kuo G, Arnaud L, Kronstad-O'Brien P, Cooper JA (September 2005). "Absence of Fyn and Src causes a reeler-like phenotype". J. Neurosci. 25 (37): 8578–86. doi:10.1523/JNEUROSCI.1656-05.2005. PMID 16162939.
- Park TJ, Curran T (December 2008). "Crk and CrkL play essential overlapping roles downstream of Dab1 in the Reelin pathway". J. Neurosci. 28 (50): 13551–62. doi:10.1523/JNEUROSCI.4323-08.2008. PMC 2628718. PMID 19074029.
- Molnár Z, Blakemore C (1992). "How are thalamocortical axons guided in the reeler mouse?". Soc. Neurosci. Abstr. 18: 778.
- Molnár Z, Blakemore C (1995). "How do thalamic axons find their way to the cortex?". Trends Neurosci. 18: 389–397. doi:10.1016/0166-2236(95)93935-q. PMID 7482804.
- Molnar Z, Adams R, Goffinet AM, Blakemore C (1998). "The role of the first postmitotic cortical cells in the development of thalamocortical innervation in the reeler mouse". J. Neurosci. 18 (15): 5746–65. doi:10.1523/jneurosci.18-15-05746.1998. PMID 9671664.
- Liu Y, Fujise N, Kosaka T (1996). "Distribution of calretinin immunoreactivity in the mouse dentate gyrus. I. General description. Exp". Brain Res. 108: 389–403. doi:10.1007/bf00227262.
- Drakew A, Deller T, Heimrich B, Gebhardt C, Del Turco D, Tielsch A, Förster E, Herz J, Frotscher M (2002). "Dentate granule cells in reeler mutants and VLDLR and ApoER2 knockout mice". Exp. Neurol. 176: 12–24. doi:10.1006/exnr.2002.7918.
- Niu S, Renfro A, Quattrocchi CC, Sheldon M, D'Arcangelo G (Jan 2004). "Reelin promotes hippocampal dendrite development through the VLDLR/ApoER2-Dab1 pathway". Neuron. 41 (1): 71–84. doi:10.1016/s0896-6273(03)00819-5. PMID 14715136.
- Matsuzaki H, Minabe Y, Nakamura K, Suzuki K, Iwata Y, Sekine Y, Tsuchiya KJ, Sugihara G, Suda S, Takei N, Nakahara D, Hashimoto K, Nairn AC, Mori N, Sato K (2007). "Disruption of reelin signaling attenuates methamphetamine-induced hyperlocomotion". Eur. J. Neurosci. 25 (11): 3376–84. doi:10.1111/j.1460-9568.2007.05564.x. PMID 17553006.
- Hadj-Sahraoui N, Frederic F, Delhaye-Bouchaud N, Mariani J (1996). "Gender effect on Purkinje cell loss in the cerebellum of the heterozygous reeler mouse". J. Neurogenet. 11 (1–2): 45–58. doi:10.3109/01677069609107062. PMID 10876649.
- Ammassari-Teule M, Sgobio C, Biamonte F, Marrone C, Mercuri NB, Keller F (March 2009). "Reelin haploinsufficiency reduces the density of PV+ neurons in circumscribed regions of the striatum and selectively alters striatal-based behaviors". Psychopharmacology. 204 (3): 511–21. doi:10.1007/s00213-009-1483-x. PMID 19277610.
- Falconer, D. S. (1951). "Two new mutants, 'trembler' and 'reeler', with neurological actions in the house mouse (Mus musculus L.)". Journal of Genetics. 50 (2): 192–205. doi:10.1007/BF02996215. PMID 24539699.
- D'Arcangelo G, Miao GG, Chen SC, Soares HD, Morgan JI, Curran T (1995). "A protein related to extracellular matrix proteins deleted in the mouse mutant reeler". Nature. 374 (6524): 719–723. Bibcode:1995Natur.374..719D. doi:10.1038/374719a0. PMID 7715726.
External links
- Development of the Cerebral Cortex: III. The Reeler Mutation - by Paul J. Lombroso, M.D.