RIPK1
Receptor-interacting serine/threonine-protein kinase 1 (RIPK1) is an enzyme that in humans is encoded by the RIPK1 gene, which is located on chromosome 6.[5][6][7] This protein belongs to the Receptor Interacting Protein (RIP) kinases family, which consists of 7 members, RIPK1 being the first member of the family.[8]




RIPK1 is known to have function in a variety of cellular pathways related to both cell survival and death. In terms of cell death, RIPK1 plays a role in apoptosis and necroptosis. Some of the cell survival pathways RIPK1 participates in include NF-κB, Akt, and JNK.[9]
Structure
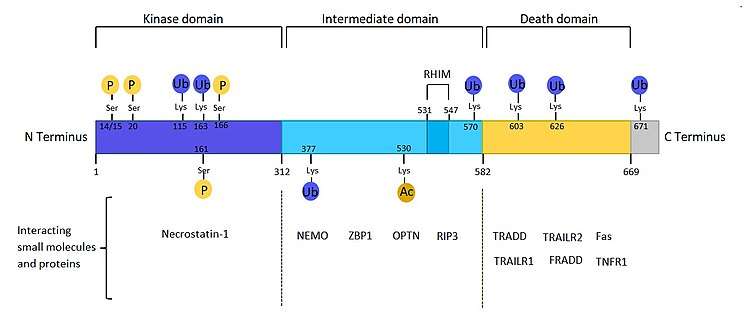
RIPK1 protein is composed of 671 amino acids, and has a molecular weight of about 76 kDa. It contains a serine/threonine kinase domain (KD) in the 300 aa N-Terminus, a death domain (DD) in the 112 aa C-Terminus, and a central region between the KD and DD called intermediate domain (ID).
- The kinase domain plays different roles in cell survival and is important in necroptosis induction. RIP interacts with TRAF2 via the kinase domain. The KD can also interact with Necrostatin-1,[10] which is an allosteric inhibitor of RIPK1 kinase activity. Overexpression of RIP lacking kinase activity can activate NF-kB.
- The death domain is homologous to the DD of other receptors such as Fas, TRAILR2 (DR5), TNFR1 and TRAILR1 (DR4), so it can bind to these receptors, as well as TRADD and FADD in the TNFR1 signalling complex. Overexpression of RIP can induce apoptosis and can activate NF-kB, but overexpression of the RIP death domain can block NF-kB activation by TNF-R1.[11]
- The intermediate domain is important for NF-kB activation and (RHIM)-dependent signalling. Via the intermediate domain, RIP can interact with TRAF2, NEMO, RIPK3, ZBP1, OPTN[12] and other small molecules and proteins, depending on cellular context.
.
Function
Although, RIPK1 has been primarily studied in the context of TNFR signaling, RIPK1 is also activated in response to diverse stimuli.[13]
The kinase domain, while important for necroptotic (programmed necrotic) functions, appears dispensable for pro-survival roles. Kinase activity of RIPK1 is also required for RIPK1-dependent apoptosis in conditions of IAP1/2 depletion, TAK1inhibition/depletion, RIPK3 depletion or MLKL depletion.[14][15] Also, proteolytic processing of RIPk1, through both caspase-dependent and -independent mechanisms, triggers lethality that is dependent on the generation of one or more specific C-terminal cleavage product(s) of RIPk1 upon stress.
Role in cell survival
It has been shown that cell survival can be regulated through different RIPK1-mediated pathways that ultimately result in the expression of NF-kB, a protein complex known to regulate transcription of DNA and thus, related to survival processes.[16]
Receptor-mediated signalling
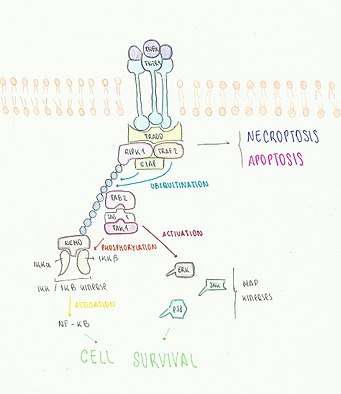
The best well-known pathway of NF-kB activation is that mediated by the death receptor TNFR1, which starts as in the necroptosis pathway with the assembly of TRADD, RIPK1, TRAF2 and clAP1 in the lipid rafts of the plasma membrane (complex I is formed). In survival signalling, RIPK1 is then polyubiquitinated, allowing NEMO (Necrosis Factor – kappa – B essential modulator) to bind to the IkB kinase or IKK complex.[17] To activate IKK, TAB2 and TAB3 adaptor proteins recruit TAK1 or MEKK3, which phosphorylate the complex. This results in the phosphorylation of the NF-kB inhibitors by the activated IKK complex, which in turn triggers their polyubiquitination and posterior degradation in the 26S proteasome.
As a result, NF-kB can now migrate to the nucleus where it will control DNA transcription by binding itself to the promoters of specific genes. Some of those genes are thought to have anti-apoptotic properties as well as to promote proteasomal degradation of RIPK1, resulting in a self-regulatory cycle.
While being in complex I, RIPK1 has also been proved to play a role in the activation of MAP (mitogen-activated protein) kinases such as JNK, ERK and p38. In particular, JNK can be found in both cell death and survival pathways, with its role in the cell death process being suppressed by activated NF-kB.[18]
Cell survival signalling can also be mediated by TLR-3 (toll-like receptors) and TLR-4. In here, RIPK1 is recruited to the receptors where it is phosphorylated and polyubiquitinated. This results in the recruit of the IKK complex activating proteins (TAK1, TAB1 and TAB2) so eventually NF-kB can now too migrate to the nucleus. RIPK2 is involved in this TLR-mediated signalling, which suggests that there might be a regulation of cell survival or death (the two possible outcomes) through the mutual interaction between the two RIPK family members.[18][19]
Genotoxic stress-mediated activation
Upon DNA damage, RIPK1 mediates another NF-kB activation pathway where two simultaneous and exclusive processes occur. A pro-apoptotic complex is created while RIPK1 also mediates the interaction between PIDD, NEMO and IKK subunits that will eventually result in the IKK complex activation after interaction with ATM kinase (a DNA double-strand breaks stimulated protein). The interaction between RIPK1 and PIDD through their death domains is thought to promote cell survival to neutralize this pro-apoptotic complex.[19]
Others
It has been observed that RIPK1 may also interact with IGF-1R (insulin-like growth factor 1 receptor) to activate JNK (c-Jun N-terminal Kinases), it may be related to epidermal growth factor receptor signalling and it is largely expressed in glioblastoma cells, suggesting that RIPK1 is indeed involved in cell survival and proliferation processes.[18]
Role in cell death
Necroptosis
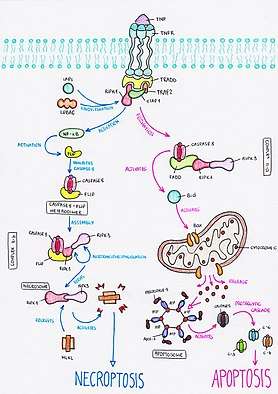
Necroptosis is a programmed form of necrosis which starts with the assembly of the TNF (tumor necrosis factor) ligand to its membrane receptor, the TNFR (tumor necrosis factor receptor). Once activated, the intracellular domain of TNFR starts the recruitment of the adaptor TNFR-1-associated death domain protein TRADD, which recruits RIPK1 and two ubiquitin ligases: TRAF2 and clAP1. This complex is called the TNFR-1 complex I.[20]
Complex-I is then modified by the IAPs (Inhibitor of Apoptosis Proteins) and the LUBAC (Linear Ubiquitination Assembly Complex), which generate linear ubiquitin linkages. The ubiquitination of complex-I leads to the activation of NF-κB , which in turn activates the expression of FLICE-like inhibitory protein FLIP. FLIP then binds to caspase-8, forming a caspase-8 FLIP heterodimer in the cytosol that disrupts the activity of caspase-8 and prevents caspase-8 mediated apoptosis from taking place.[21]
The assembly of complex II-b then starts in the cytosol. This new complex contains the caspase-8 FLIP heterodimer as well as RIPK1 and RIPK3. Caspase inhibition within this complex allows RIPK1 and RIPK3 to autotransphosphorylate each other, forming another complex called the necrosome.[22] The necrosome starts recruiting MLKL (Mixed Kinase Domain Like protein), which is phosphorylated by RIPK3 and immediately translocates to lipid rafts inside the plasma membrane. This leads to the formation of pores in the membrane, allowing the sodium influx to increase -and consequently the osmotic pressure-, which eventually causes cell membrane rupture.[22]
Apoptosis
The apoptotic extrinsic pathway starts with the formation of the TNFR-1 complex-I, which contains TRADD, RIPK1, and two ubiquitin ligases:TRAF2 and clAP1.[23][20]
Unlike the necroptotic pathway, this pathway doesn’t include the inhibition of caspase-8. Thus, in absence of NF-κB function, FLIP is not produced, and therefore active caspase-8 assembles with FADD, RIPK1 and RIPK3 in the cytosol, forming what is known as complex IIa.[22]
Caspase-8 activates Bid, a protein that binds to the mitochondrial membrane, allowing the release of intermembrane mitochondrial molecules such as cytochrome c. Cytochrome c then assembles with Apaf 1 and ATP molecules, forming a complex called apoptosome. The activation of caspase 3 and 9 by the apoptosome starts a proteolitic cascade that eventually leads to the degradation of organelles and proteins, and the fragmentation of the DNA, inducing apoptotic cell death.
Neurodegenerative diseases
Alzheimer's disease
Patients with Alzheimer's disease, a neurodegenerative disease characterized by a cognitive deterioration and a behavioural disorder, experience a chronic brain inflammation which leads to the atrophy of several brain regions.
A sign of this inflammation is an increased number of microglia, a type of glial cells located in the brain and the spinal cord. RIPK1 is known to appear in larger quantities in brains from those affected with AD.[24] This enzyme regulates not only necroptosis, but cell inflammation as well, and as a result it is involved in the regulation of microglial functions, specially those associated with the appearance and development of neurodegenerative diseases such as AD.[24]
Amyotrophic Lateral Sclerosis
Amyotrophic Lateral Sclerosis (ALS) is characterized by the degeneration of motor neurons which leads to the progressive loss of mobility. Consequently, patients are unable to do any physical activity due to the atrophy of their muscles.[25]
The optineurin gene (OPTN) and its mutation are known to be involved in ALS. When the organism loses OPTN, the dysmyelination of axons and its degeneration start. The degeneration of the axons is produced by several components from the Central Nervous System (CNS) including RIPK1 and another enzyme from the Receptor Interacting Protein kinases family, RIPK3, as well as other proteins such as MLKL.[26]
Once RIPK1, RIPK3 and MLKL have contributed to the dysmyelination and the consequent degeneration of axons, the nerve impulse can't to go from one neuron to another due to the lack of myelin, which leads to the consequent mobility problems as the nerve impulse does not arrive to its final destination.[27]
Autoinflamatory disease
An autoinflammatory disease characterised by recurrent fevers and lymphadenopathy has been associated with mutations in this gene.[28]
Interactions
RIPK1 has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000137275 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000021408 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Stanger BZ, Leder P, Lee TH, Kim E, Seed B (May 1995). "RIP: a novel protein containing a death domain that interacts with Fas/APO-1 (CD95) in yeast and causes cell death". Cell. 81 (4): 513–23. doi:10.1016/0092-8674(95)90072-1. PMID 7538908.
- Hsu H, Huang J, Shu HB, Baichwal V, Goeddel DV (April 1996). "TNF-dependent recruitment of the protein kinase RIP to the TNF receptor-1 signaling complex". Immunity. 4 (4): 387–96. doi:10.1016/S1074-7613(00)80252-6. PMID 8612133.
- "Entrez Gene: RIPK1 receptor (TNFRSF)-interacting serine-threonine kinase 1".
- Festjens N, Vanden Berghe T, Cornelis S, Vandenabeele P (March 2007). "RIP1, a kinase on the crossroads of a cell's decision to live or die". Cell Death and Differentiation. 14 (3): 400–10. doi:10.1038/sj.cdd.4402085. PMID 17301840.
- Lin, Yong (2014). "RIP1-Mediated Signaling Pathways in Cell Survival and Death Control". Necrotic Cell Death. Springer New York. pp. 23–43. doi:10.1007/978-1-4614-8220-8_2. ISBN 9781461482192.
- Vandenabeele P, Grootjans S, Callewaert N, Takahashi N (February 2013). "Necrostatin-1 blocks both RIPK1 and IDO: consequences for the study of cell death in experimental disease models". Cell Death and Differentiation. 20 (2): 185–7. doi:10.1038/cdd.2012.151. PMC 3554339. PMID 23197293.
- Online Mendelian Inheritance in Man (OMIM): Receptor-interacting serine/threonine kinase 1 (RIPK1) - 603453
- Zhu G, Wu CJ, Zhao Y, Ashwell JD (August 2007). "Optineurin negatively regulates TNFalpha- induced NF-kappaB activation by competing with NEMO for ubiquitinated RIP". Current Biology. 17 (16): 1438–43. doi:10.1016/j.cub.2007.07.041. PMID 17702576.
- Vanlangenakker N, Vanden Berghe T, Vandenabeele P (January 2012). "Many stimuli pull the necrotic trigger, an overview". Cell Death and Differentiation. 19 (1): 75–86. doi:10.1038/cdd.2011.164. PMC 3252835. PMID 22075985.
- Dondelinger Y, Aguileta MA, Goossens V, Dubuisson C, Grootjans S, Dejardin E, Vandenabeele P, Bertrand MJ (October 2013). "RIPK3 contributes to TNFR1-mediated RIPK1 kinase-dependent apoptosis in conditions of cIAP1/2 depletion or TAK1 kinase inhibition". Cell Death and Differentiation. 20 (10): 1381–92. doi:10.1038/cdd.2013.94. PMC 3770330. PMID 23892367.
- Remijsen Q, Goossens V, Grootjans S, Van den Haute C, Vanlangenakker N, Dondelinger Y, Roelandt R, Bruggeman I, Goncalves A, Bertrand MJ, Baekelandt V, Takahashi N, Berghe TV, Vandenabeele P (January 2014). "Depletion of RIPK3 or MLKL blocks TNF-driven necroptosis and switches towards a delayed RIPK1 kinase-dependent apoptosis". Cell Death & Disease. 5: e1004. doi:10.1038/cddis.2013.531. PMC 4040672. PMID 24434512.
- Christofferson DE, Li Y, Yuan J (2014). "Control of life-or-death decisions by RIP1 kinase". Annual Review of Physiology. 76: 129–50. doi:10.1146/annurev-physiol-021113-170259. PMID 24079414.
- Zhang J, Zhang H, Li J, Rosenberg S, Zhang EC, Zhou X, Qin F, Farabaugh M (2011). "RIP1-mediated regulation of lymphocyte survival and death responses". Immunologic Research. 51 (2–3): 227–36. doi:10.1007/s12026-011-8249-3. PMC 3244575. PMID 22038529.
- Lin Y (2014). "RIP1-Mediated Signaling Pathways in Cell Survival and Death Control". Necrotic Cell Death. Springer New York. pp. 23–43. doi:10.1007/978-1-4614-8220-8_2. ISBN 9781461482192.
- Festjens N, Vanden Berghe T, Cornelis S, Vandenabeele P (2007). "RIP1, a kinase on the crossroads of a cell's decision to live or die". Cell Death and Differentiation. 14 (3): 400–10. doi:10.1038/sj.cdd.4402085. PMID 17301840.
- Hsu H, Huang J, Shu HB, Baichwal V, Goeddel DV (April 1996). "TNF-dependent recruitment of the protein kinase RIP to the TNF receptor-1 signaling complex". Immunity. 4 (4): 387–96. doi:10.1016/s1074-7613(00)80252-6. PMID 8612133.
- Linkermann A, Green DR (January 2014). "Necroptosis". The New England Journal of Medicine. 370 (5): 455–65. doi:10.1056/NEJMra1310050. PMC 4035222. PMID 24476434.
- Cho YS, Challa S, Moquin D, Genga R, Ray TD, Guildford M, Chan FK (June 2009). "Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation". Cell. 137 (6): 1112–23. doi:10.1016/j.cell.2009.05.037. PMC 2727676. PMID 19524513.
- Green DR, Oberst A, Dillon CP, Weinlich R, Salvesen GS (October 2011). "RIPK-dependent necrosis and its regulation by caspases: a mystery in five acts". Molecular Cell. 44 (1): 9–16. doi:10.1016/j.molcel.2011.09.003. PMC 3192321. PMID 21981915.
- Ofengeim D, Mazzitelli S, Ito Y, DeWitt JP, Mifflin L, Zou C, Das S, Adiconis X, Chen H, Zhu H, Kelliher MA, Levin JZ, Yuan J (September 2017). "RIPK1 mediates a disease-associated microglial response in Alzheimer's disease". Proceedings of the National Academy of Sciences of the United States of America. 114 (41): E8788–E8797. doi:10.1073/pnas.1714175114. PMC 5642727. PMID 28904096.
- "What is ALS?". ALSA.org. Retrieved 2017-10-21.
- Ito Y, Ofengeim D, Najafov A, Das S, Saberi S, Li Y, Hitomi J, Zhu H, Chen H, Mayo L, Geng J, Amin P, DeWitt JP, Mookhtiar AK, Florez M, Ouchida AT, Fan JB, Pasparakis M, Kelliher MA, Ravits J, Yuan J (August 2016). "RIPK1 mediates axonal degeneration by promoting inflammation and necroptosis in ALS". Science. 353 (6299): 603–8. doi:10.1126/science.aaf6803. PMC 5444917. PMID 27493188.
- "Transmission of Nerve Impulses". www.cliffsnotes.com. Retrieved 2017-10-21.
- Tao P, Sun J, Wu Z, Wang S, Wang J, Li W, et al. (December 2019). "A dominant autoinflammatory disease caused by non-cleavable variants of RIPK1". Nature. doi:10.1038/s41586-019-1830-y. PMID 31827280.
- Bertrand MJ, Milutinovic S, Dickson KM, Ho WC, Boudreault A, Durkin J, Gillard JW, Jaquith JB, Morris SJ, Barker PA (June 2008). "cIAP1 and cIAP2 facilitate cancer cell survival by functioning as E3 ligases that promote RIP1 ubiquitination". Molecular Cell. 30 (6): 689–700. doi:10.1016/j.molcel.2008.05.014. PMID 18570872.
- Liao W, Xiao Q, Tchikov V, Fujita K, Yang W, Wincovitch S, Garfield S, Conze D, El-Deiry WS, Schütze S, Srinivasula SM (May 2008). "CARP-2 is an endosome-associated ubiquitin ligase for RIP and regulates TNF-induced NF-kappaB activation". Current Biology. 18 (9): 641–9. doi:10.1016/j.cub.2008.04.017. PMC 2587165. PMID 18450452.
- Chaudhary PM, Eby MT, Jasmin A, Kumar A, Liu L, Hood L (September 2000). "Activation of the NF-kappaB pathway by caspase 8 and its homologs". Oncogene. 19 (39): 4451–60. doi:10.1038/sj.onc.1203812. PMID 11002417.
- Oshima S, Turer EE, Callahan JA, Chai S, Advincula R, Barrera J, Shifrin N, Lee B, Benedict Yen TS, Yen B, Woo T, Malynn BA, Ma A (February 2009). "ABIN-1 is a ubiquitin sensor that restricts cell death and sustains embryonic development". Nature. 457 (7231): 906–9. doi:10.1038/nature07575. PMC 2642523. PMID 19060883.
- Kataoka T, Budd RC, Holler N, Thome M, Martinon F, Irmler M, Burns K, Hahne M, Kennedy N, Kovacsovics M, Tschopp J (June 2000). "The caspase-8 inhibitor FLIP promotes activation of NF-kappaB and Erk signaling pathways". Current Biology. 10 (11): 640–8. doi:10.1016/S0960-9822(00)00512-1. PMID 10837247.
- Duan H, Dixit VM (January 1997). "RAIDD is a new 'death' adaptor molecule" (PDF). Nature. 385 (6611): 86–9. doi:10.1038/385086a0. hdl:2027.42/62739. PMID 8985253.
- Ahmad M, Srinivasula SM, Wang L, Talanian RV, Litwack G, Fernandes-Alnemri T, Alnemri ES (February 1997). "CRADD, a novel human apoptotic adaptor molecule for caspase-2, and FasL/tumor necrosis factor receptor-interacting protein RIP". Cancer Research. 57 (4): 615–9. PMID 9044836.
- Yu PW, Huang BC, Shen M, Quast J, Chan E, Xu X, Nolan GP, Payan DG, Luo Y (May 1999). "Identification of RIP3, a RIP-like kinase that activates apoptosis and NFkappaB". Current Biology. 9 (10): 539–42. doi:10.1016/S0960-9822(99)80239-5. PMID 10339433.
- Li J, McQuade T, Siemer AB, Napetschnig J, Moriwaki K, Hsiao YS, Damko E, Moquin D, Walz T, McDermott A, Chan FK, Wu H (July 2012). "The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis". Cell. 150 (2): 339–50. doi:10.1016/j.cell.2012.06.019. PMC 3664196. PMID 22817896.
- Shembade N, Parvatiyar K, Harhaj NS, Harhaj EW (March 2009). "The ubiquitin-editing enzyme A20 requires RNF11 to downregulate NF-kappaB signalling". The EMBO Journal. 28 (5): 513–22. doi:10.1038/emboj.2008.285. PMC 2657574. PMID 19131965.
- Chen D, Li X, Zhai Z, Shu HB (May 2002). "A novel zinc finger protein interacts with receptor-interacting protein (RIP) and inhibits tumor necrosis factor (TNF)- and IL1-induced NF-kappa B activation". The Journal of Biological Chemistry. 277 (18): 15985–91. doi:10.1074/jbc.M108675200. PMID 11854271.
- Sanz L, Sanchez P, Lallena MJ, Diaz-Meco MT, Moscat J (June 1999). "The interaction of p62 with RIP links the atypical PKCs to NF-kappaB activation". The EMBO Journal. 18 (11): 3044–53. doi:10.1093/emboj/18.11.3044. PMC 1171386. PMID 10356400.
- Kim JW, Choi EJ, Joe CO (September 2000). "Activation of death-inducing signaling complex (DISC) by pro-apoptotic C-terminal fragment of RIP". Oncogene. 19 (39): 4491–9. doi:10.1038/sj.onc.1203796. PMID 11002422.
- Blankenship JW, Varfolomeev E, Goncharov T, Fedorova AV, Kirkpatrick DS, Izrael-Tomasevic A, Phu L, Arnott D, Aghajan M, Zobel K, Bazan JF, Fairbrother WJ, Deshayes K, Vucic D (January 2009). "Ubiquitin binding modulates IAP antagonist-stimulated proteasomal degradation of c-IAP1 and c-IAP2(1)". The Biochemical Journal. 417 (1): 149–60. doi:10.1042/BJ20081885. PMID 18939944.
- Newton K, Matsumoto ML, Wertz IE, Kirkpatrick DS, Lill JR, Tan J, Dugger D, Gordon N, Sidhu SS, Fellouse FA, Komuves L, French DM, Ferrando RE, Lam C, Compaan D, Yu C, Bosanac I, Hymowitz SG, Kelley RF, Dixit VM (August 2008). "Ubiquitin chain editing revealed by polyubiquitin linkage-specific antibodies". Cell. 134 (4): 668–78. doi:10.1016/j.cell.2008.07.039. PMID 18724939.
- Varfolomeev E, Goncharov T, Fedorova AV, Dynek JN, Zobel K, Deshayes K, Fairbrother WJ, Vucic D (September 2008). "c-IAP1 and c-IAP2 are critical mediators of tumor necrosis factor alpha (TNFalpha)-induced NF-kappaB activation". The Journal of Biological Chemistry. 283 (36): 24295–9. doi:10.1074/jbc.C800128200. PMC 3259840. PMID 18621737.
- Takeuchi M, Rothe M, Goeddel DV (August 1996). "Anatomy of TRAF2. Distinct domains for nuclear factor-kappaB activation and association with tumor necrosis factor signaling proteins". The Journal of Biological Chemistry. 271 (33): 19935–42. doi:10.1074/jbc.271.33.19935. PMID 8702708.
- Tada K, Okazaki T, Sakon S, Kobarai T, Kurosawa K, Yamaoka S, Hashimoto H, Mak TW, Yagita H, Okumura K, Yeh WC, Nakano H (September 2001). "Critical roles of TRAF2 and TRAF5 in tumor necrosis factor-induced NF-kappa B activation and protection from cell death". The Journal of Biological Chemistry. 276 (39): 36530–4. doi:10.1074/jbc.M104837200. PMID 11479302.
- Malinin NL, Boldin MP, Kovalenko AV, Wallach D (February 1997). "MAP3K-related kinase involved in NF-kappaB induction by TNF, CD95 and IL-1". Nature. 385 (6616): 540–4. doi:10.1038/385540a0. PMID 9020361.
- Ma Q, Zhou L, Shi H, Huo K (June 2008). "NUMBL interacts with TAB2 and inhibits TNFalpha and IL-1beta-induced NF-kappaB activation". Cellular Signalling. 20 (6): 1044–51. doi:10.1016/j.cellsig.2008.01.015. PMID 18299187.
- Belizário J, Vieira-Cordeiro L, Enns S (2015). "Necroptotic Cell Death Signaling and Execution Pathway: Lessons from Knockout Mice". Mediators of Inflammation. 2015: 128076. doi:10.1155/2015/128076. PMC 4600508. PMID 26491219.
Further reading
- Duan H, Dixit VM (January 1997). "RAIDD is a new 'death' adaptor molecule" (PDF). Nature. 385 (6611): 86–9. doi:10.1038/385086a0. hdl:2027.42/62739. PMID 8985253.
- Chaudhary PM, Eby M, Jasmin A, Bookwalter A, Murray J, Hood L (December 1997). "Death receptor 5, a new member of the TNFR family, and DR4 induce FADD-dependent apoptosis and activate the NF-kappaB pathway". Immunity. 7 (6): 821–30. doi:10.1016/S1074-7613(00)80400-8. PMID 9430227.
- Juo P, Kuo CJ, Yuan J, Blenis J (September 1998). "Essential requirement for caspase-8/FLICE in the initiation of the Fas-induced apoptotic cascade". Current Biology. 8 (18): 1001–8. doi:10.1016/S0960-9822(07)00420-4. PMID 9740801.
- Li Y, Kang J, Friedman J, Tarassishin L, Ye J, Kovalenko A, Wallach D, Horwitz MS (February 1999). "Identification of a cell protein (FIP-3) as a modulator of NF-kappaB activity and as a target of an adenovirus inhibitor of tumor necrosis factor alpha-induced apoptosis". Proceedings of the National Academy of Sciences of the United States of America. 96 (3): 1042–7. doi:10.1073/pnas.96.3.1042. PMC 15347. PMID 9927690.
- Sun X, Lee J, Navas T, Baldwin DT, Stewart TA, Dixit VM (June 1999). "RIP3, a novel apoptosis-inducing kinase". The Journal of Biological Chemistry. 274 (24): 16871–5. doi:10.1074/jbc.274.24.16871. PMID 10358032.
- Izumi KM, Cahir McFarland ED, Ting AT, Riley EA, Seed B, Kieff ED (August 1999). "The Epstein-Barr virus oncoprotein latent membrane protein 1 engages the tumor necrosis factor receptor-associated proteins TRADD and receptor-interacting protein (RIP) but does not induce apoptosis or require RIP for NF-kappaB activation". Molecular and Cellular Biology. 19 (8): 5759–67. doi:10.1128/mcb.19.8.5759. PMC 84426. PMID 10409763.
- Lin Y, Devin A, Rodriguez Y, Liu ZG (October 1999). "Cleavage of the death domain kinase RIP by caspase-8 prompts TNF-induced apoptosis". Genes & Development. 13 (19): 2514–26. doi:10.1101/gad.13.19.2514. PMC 317073. PMID 10521396.
- Chaudhary PM, Jasmin A, Eby MT, Hood L (October 1999). "Modulation of the NF-kappa B pathway by virally encoded death effector domains-containing proteins". Oncogene. 18 (42): 5738–46. doi:10.1038/sj.onc.1202976. PMID 10523854.
- Lewis J, Devin A, Miller A, Lin Y, Rodriguez Y, Neckers L, Liu ZG (April 2000). "Disruption of hsp90 function results in degradation of the death domain kinase, receptor-interacting protein (RIP), and blockage of tumor necrosis factor-induced nuclear factor-kappaB activation". The Journal of Biological Chemistry. 275 (14): 10519–26. doi:10.1074/jbc.275.14.10519. PMID 10744744.
- Inohara N, Koseki T, Lin J, del Peso L, Lucas PC, Chen FF, Ogura Y, Núñez G (September 2000). "An induced proximity model for NF-kappa B activation in the Nod1/RICK and RIP signaling pathways". The Journal of Biological Chemistry. 275 (36): 27823–31. doi:10.1074/jbc.M003415200. PMID 10880512.