Photosynthetic picoplankton
Photosynthetic picoplankton or picophytoplankton is the fraction of the phytoplankton performing photosynthesis composed by cells between 0.2 and 2 µm (picoplankton). It is especially important in the central oligotrophic regions of the world oceans that have very low concentration of nutrients.

History
- 1952: Description of the first truly picoplanktonic species, Chromulina pusilla, by Butcher.[1] This species was renamed in 1960 to Micromonas pusilla[2] and a few studies have found it to be abundant in temperate oceanic waters, although very little such quantification data exists for eukaryotic picophytoplankton.
- 1979: Discovery of marine Synechococcus by Waterbury[3] and confirmation with electron microscopy by Johnson and Sieburth.[4]
- 1982: The same Johnson and Sieburth demonstrate the importance of small eukaryotes by electron microscopy.[5]
- 1983: W.K.W Li and colleagues, including Trevor Platt show that a large fraction of marine primary production is due to organisms smaller than 2 µm.[6]
- 1986: Discovery of "prochlorophytes" by Chisholm and Olson in the Sargasso Sea,[7] named in 1992 as Prochlorococcus marinus.[8]
- 1994: Discovery in the Thau lagoon in France of the smallest photosynthetic eukaryote known to date, Ostreococcus tauri, by Courties.[9]
- 2001: Through sequencing of the ribosomal RNA gene extracted from marine samples, several European teams discover that eukaryotic picoplankton are highly diverse.[10][11] This finding followed on the first discovery of such eukaryotic diversity in 1998 by Rappe and colleagues at Oregon State University, who were the first to apply rRNA sequencing to eukaryotic plankton in the open-ocean, where they discovered sequences that seemed distant from known phytoplankton[12] The cells containing DNA matching one of these novel sequences were recently visualized and further analyzed using specific probes and found to be broadly distributed.[13]
Methods of study
Because of its very small size, picoplankton is difficult to study by classic methods such as optical microscopy. More sophisticated methods are needed.
- Epifluorescence microscopy allows researchers to detect certain groups of cells possessing fluorescent pigments such as Synechococcus which possess phycoerythrin.
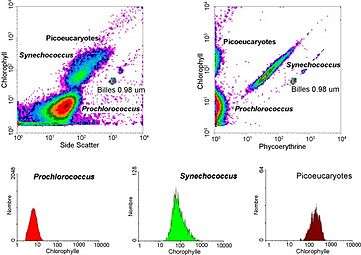
- Flow cytometry measures the size (" forward scatter ") and fluorescence of 1,000 in 10,000 cells per second. It allows one to determine very easily the concentration of the various picoplankton populations on marine samples. Three groups of cells (Prochlorococcus, Synechococcus and picoeukaryotes) can be distinguished. For example Synechococcus is characterized by the double fluorescence of its pigments: orange for phycoerythrin and red for chlorophyll. Flow cytometry also allows researchers to sort out specific populations (for example Synechococcus) in order put them in culture, or to make more detailed analyses.
- Analysis of photosynthetic pigments such as chlorophyll or carotenoids by high precision chromatography (HPLC) allows researchers to determine the various groups of algae present in a sample.
- Molecular biology techniques:
- Cloning and sequencing of genes such as that of ribosomal RNA, which allows researchers to determine total diversity within a sample
- DGGE (Denaturing Gel Electrophoresis), that is faster than the previous approach allows researchers to have an idea of the global diversity within a sample
- In situ hybridization (FISH) uses fluorescent probes recognizing specific taxon, for example a species, a genus or a class.[14] This original description as a species is now thought to be composed of a number of different cryptic species, a finding that has been confirmed by a genome sequencing project of two strains led by researchers at the Monterey Bay Aquarium Research Institute.[15]
- Quantitative PCR can be used, as FISH, to determine, the abundance of specific groups. It has the main advantage to allow the rapid analysis of a large number of samples simultaneously,[16] but requires more sophisticated controls and calibrations.
Composition
Three major groups of organisms constitute photosynthetic picoplankton...
- Cyanobacteria belonging to the genus Synechococcus of a size of 1 µm (micrometer) were first discovered in 1979 by J. Waterbury[3] (Woods Hole Oceanographic Institution). They are quite ubiquitous, but most abundant in relatively mesotrophic waters.
- Cyanobacteria belonging to the genus Prochlorococcus are particularly remarkable. With a typical size of 0.6 µm, Prochlorococcus was discovered only in 1988[7] by two American researchers, Sallie W. (Penny) Chisholm (Massachusetts Institute of Technology) and R.J. Olson (Woods Hole Oceanographic Institution). In spite of its small size, this photosynthetic organism is undoubtedly the most abundant of the planet: indeed its density can reach up to 100 million cells per liter and it can be found down to a depth of 150 m in all the intertropical belt.[17]
- Picoplanktonic eukaryotes are the least well known, as demonstrated by the recent discovery of major groups. Andersen created in 1993 a new class of brown algae, the Pelagophyceae.[18] More surprising still, the discovery in 1994[9] of a eukaryote of very small size, Ostreococcus tauri, dominating the phytoplanktonic biomass of a French brackish lagoon (étang de Thau), shows that these organisms can also play a major ecological role in coastal environments. In 1999, yet a new class of alga was discovered,[19] the Bolidophyceae, very close genetically of diatoms, but quite different morphologically. At the present time, about 50 species are known belonging to several classes.
Algal classes containing picoplankton species Classes Picoplanktonic genera Chlorophyceae Nannochloris Prasinophyceae Micromonas, Ostreococcus, Pycnococcus Prymnesiophyceae Imantonia Pelagophyceae Pelagomonas Bolidophyceae Bolidomonas Dictyochophyceae Florenciella
The use of molecular approaches implemented since the 1990s for bacteria, were applied to the photosynthetic picoeukaryotes only 10 years later around 2000. They revealed a very wide diversity[10][11] and brought to light the importance of the following groups in the picoplankton :
In temperate coastal environment, the genus Micromonas (Prasinophyceae) seems dominant.[14] However, in numerous oceanic environments, the dominant species of eukaryotic picoplankton remain still unknown.[20]
Ecology
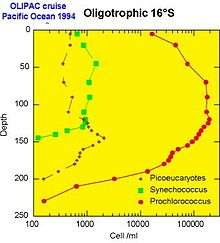
Each picoplanktonic population occupies a specific ecological niche in the oceanic environment.
- The Synechococcus cyanobacterium is generally abundant in mesotrophic environments, such as near the equatorial upwelling or in coastal regions.
- The Prochlorococcus cyanobacterium replaces it when the waters becomes impoverished in nutrients (i.e., oligotrophic). On the other hand, in temperate regions such as the North Atlantic Ocean, Prochlorococcus is absent because the cold waters prevent its development.
- The diversity of eukaryotes derives from their presence in a large variety of environments. In oceanic regions, they are often observed at depth, at the base of the well-lit layer (the "euphotic" layer). In coastal regions, certain sorts of picoeukaryotes such as Micromonas dominate. As with larger plankton, their abundance follows a seasonal cycle with a maximum in summer.
Thirty years ago, it was hypothesized that the speed of division for micro-organisms in central oceanic ecosystems was very slow, of the order of one week or one month per generation. This hypothesis was supported by the fact that the biomass (estimated for example by the contents of chlorophyll) was very stable over time. However, with the discovery of the picoplankton, it was found that the system was much more dynamic than previously thought. In particular, small predators of a size of a few micrometres which ingest picoplanktonic algae as quickly as they were produced were found to be ubiquitous. This extremely sophisticated predator-prey system is nearly always at equilibrium and results in a quasi-constant picoplankton biomass. This close equivalence between production and consumption makes it extremely difficult to measure precisely the speed at which the system turns over.
In 1988, two American researchers, Carpenter and Chang, suggested estimating the speed of cell division of phytoplankton by following the course of DNA replication by microscopy. By replacing the microscope by a flow cytometer, it is possible to follow the DNA content of picoplankton cells over time. This allowed researchers to establish that picoplankton cells are highly synchronous: they replicate their DNA and then divide all at the same time at the end of the day. This synchronization could be due to the presence of an internal biological clock.
Genomics
In the 2000s, genomics allowed to cross a supplementary stage. Genomics consists in determining the complete sequence of genome of an organism and to list every gene present. It is then possible to get an idea of the metabolic capacities of the targeted organisms and understand how it adapts to its environment. To date, the genomes of several types of Prochlorococcus[21][22] and Synechococcus,[23] and of a strain of Ostreococcus[24] have been determined. The complete genomes of two different Micromonas strains revealed that they were quite different (different species) - and had similarities with land plants.[15] Several other cyanobacteria and of small eukaryotes (Bathycoccus, Pelagomonas) are under sequencing. In parallel, genome analyses begin to be done directly from oceanic samples (ecogenomics or métagenomics),[25] allowing us to access to large sets of gene for uncultivated organisms.
Genomes of photosynthetic picoplankton strains
that have been sequenced to dateGenus Strain Sequencing center Remark Prochlorococcus MED4 JGI SS120 Genoscope MIT9312 JGI MIT9313 JGI NATL2A JGI CC9605 JGI CC9901 JGI Synechococcus WH8102 JGI WH7803 Genoscope RCC307 Génoscope CC9311 TIGR [26] Ostreococcus OTTH95 Genoscope Micromonas RCC299 and CCMP1545 JGI [15]
See also
Notes and references
- Butcher, R. (1952). Contributions to our knowledge of the smaller marine algae. Journal of the Marine Biological Association of the UK 31: 175-91.
- Manton, I. & Parke, M. (1960). Further observations on small green flagellates with special reference to possible relatives of Chromulina pusilla Butcher. Journal of the Marine Biological Association of the UK 39: 275-98.
- Waterbury, J. B. et al. (1979). Wide-spread occurrence of a unicellular, marine planktonic, cyanobacterium. Nature 277: 293-4.
- Johnson, P. W. & Sieburth, J. M. (1979). Chroococcoid cyanobacteria in the sea: a ubiquitous and diverse phototrophic biomass. Limnology and Oceanography 24: 928-35.
- Johnson, P. W. & Sieburth, J. M. (1982). In-situ morphology and occurrence of eucaryotic phototrophs of bacterial size in the picoplankton of estuarine and oceanic waters. Journal of Phycology 18: 318-27.
- Li, W. K. W. et al. (1983). Autotrophic picoplankton in the tropical ocean. Science 219: 292-5.
- Chisholm, S. W. et al. (1988). A novel free-living prochlorophyte occurs at high cell concentrations in the oceanic euphotic zone. Nature 334: 340-3.
- Chisholm, S. W. et al. (1992). Prochlorococcus marinus nov. gen. nov. sp.: an oxyphototrophic marine prokaryote containing divinyl chlorophyll a and b. Archives of Microbiology 157: 297-300.
- Courties, C. et al. (1994). Smallest eukaryotic organism. Nature 370: 255.
- López-García, P. et al. (2001). Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton. Nature 409: 603-7.
- Moon-van der Staay, S. Y. et al. (2001). Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity. Nature 409: 607-10.
- Rappe, M. et al. (1998). Phylogenetic diversity of ultraplankton plastid Small-Subunit rRNA genes recovered in environmental nucleic acid samples from the Pacific and Atlantic coasts of the United States. Applied and Environmental Microbiology 64294-303.
- Kim, E., Harrison, J., Sudek, S. et al. (2011). Newly identified and diverse plastid-bearing branch on the eukaryotic tree of life. Proceedings of the National Academy of Sciences USA. Vol. 108: 1496-1500.
- Not, F. et al. (2004). A single species Micromonas pusilla (Prasinophyceae) dominates the eukaryotic picoplankton in the western English Channel. Applied and Environmental Microbiology 70: 4064-72.
- Worden, A.Z., et al. (2009). Green evolution and dynamic adaptations revealed by genomes of the marine picoeukaryotes Micromonas. Science 324: 268-272.
- Johnson, Z. I. et al. (2006). Niche partitioning among Prochlorococcus ecotypes along ocean-scale environmental gradients. Science 311: 1737-40.
- Partensky, F. et al. (1999). Prochlorococcus, a marine photosynthetic prokaryote of global significance. Microbiology and Molecular Biology Reviews 63: 106-27.
- Andersen, R. A. et al. (1993). Ultrastructure and 18S rRNA gene sequence for Pelagomonas calceolata gen. and sp. nov. and the description of a new algal class, the Pelagophyceae classis nov. Journal of Phycology 29: 701-15.
- Guillou, L. et al. (1999). Bolidomonas: a new genus with two species belonging to a new algal class, the Bolidophyceae (Heterokonta). Journal of Phycology 35: 368-81.
- Worden, A.Z. & Not, F.(2008) Ecology and Diversity of Picoeukaryotes. Book Chapter in: Microbial Ecology of the Ocean, 2nd Edition. Ed. D. Kirchman. Wiley.
- Rocap, G. et al. (2003). Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation. Nature 424: 1042-7.
- Dufresne, A. et al. (2003). Genome sequence of the cyanobacterium Prochlorococcus marinus SS120, a nearly minimal oxyphototrophic genome. Proceedings of the National Academy of Sciences of the United States of America 100: 10020-5.
- Palenik, B. et al. (2003). The genome of a motile marine Synechococcus. Nature 424: 1037-42.
- Derelle, E. et al. (2006). Genome analysis of the smallest free-living eukaryote Ostreococcus tauri unveils many unique features. Proceedings of the National Academy of Sciences of the United States of America 103: 11647-52.
- Venter, J. C. et al. (2004). Environmental genome shotgun sequencing of the Sargasso Sea. Science 304: 66-74.
- Palenik, B. et al. (2006). Genome sequence of Synechococcus CC9311: Insights into adaptation to a coastal environment. PNAS 103: 13555-9.
Bibliography
- Cyanobacteria
- Zehr, J. P., Waterbury, J. B., Turner, P. J., Montoya, J. P., Omoregie, E., Steward, G. F., Hansen, A. & Karl, D. M. 2001. Unicellular cyanobacteria fix N2 in the subtropical North Pacific Ocean. Nature 412:635-8
- Eukaryotes
- Butcher, R. 1952. Contributions to our knowledge of the smaller marine algae. J. Mar. Biol. Assoc. UK. 31:175-91.
- Manton, I. & Parke, M. 1960. Further observations on small green flagellates with special reference to possible relatives of Chromulina pusilla Butcher. J. Mar. Biol. Assoc. UK. 39:275-98.
- Eikrem, W., Throndsen, J. 1990. The ultrastructure of Bathycoccus gen. nov. and B. prasinos sp. nov., a non-motile picoplanktonic alga (Chlorophyta, Prasinophyceae) from the Mediterranean and Atlantic. Phycologia 29:344-350
- Chrétiennot-Dinet, M. J., Courties, C., Vaquer, A., Neveux, J., Claustre, H., et al. 1995. A new marine picoeucaryote: Ostreococcus tauri gen et sp nov (Chlorophyta, Prasinophyceae). Phycologia 34:285-292
- Sieburth, J. M., M. D. Keller, P. W. Johnson, and S. M. Myklestad. 1999. Widespread occurrence of the oceanic ultraplankter, Prasinococcus capsulatus (Prasinophyceae), the diagnostic "Golgi-decapore complex" and the newly described polysaccharide "capsulan". J. Phycol. 35: 1032-1043.
- Not, F., Valentin, K., Romari, K., Lovejoy, C., Massana, R., Töbe, K., Vaulot, D. & Medlin, L. K. 2007. Picobiliphytes, a new marine picoplanktonic algal group with unknown affinities to other eukaryotes. Science 315:252-4.
- Vaulot, D., Eikrem, W., Viprey, M. & Moreau, H. 2008. The diversity of small eukaryotic phytoplankton (≤3 μm) in marine ecosystems. FEMS Microbiol. Rev. 32:795-820.
- Ecology
- Platt, T., Subba-Rao, D. V. & Irwin, B. 1983. Photosynthesis of picoplankton in the oligotrophic ocean. Nature 300:701-4.
- Stomp M, Huisman J, de Jongh F, Veraart AJ, Gerla D, Rijkeboer M, Ibelings BW, Wollenzien UIA, Stal LJ. 2004. Adaptive divergence in pigment composition promotes phytoplankton biodiversity. Nature 432: 104-107.
- Campbell, L., Nolla, H. A. & Vaulot, D. 1994. The importance of Prochlorococcus to community structure in the central North Pacific Ocean. Limnol. Oceanogr. 39:954-61.
- Molecular Biology and Genomes
- Rappé, M. S., P. F. Kemp, and S. J. Giovannoni. 1995. Chromophyte plastid 16S ribosomal RNA genes found in a clone library from Atlantic Ocean seawater. J. Phycol. 31: 979-988.