Notonecta glauca
Notonecta glauca (common backswimmer) is a species of aquatic insect, and a type of backswimmer. This species is found in large parts of Europe, North Africa, and east through Asia to Siberia and China.[1] In much of its range it is the most common backswimmer species.[2] It is also the most widespread and abundant of the four British backswimmers.[3] Notonecta glauca are Hemiptera (true bug) predators,[2] that are approximately 13–16 mm in length.[4] Females have a larger body size compared to males.[2] These water insects swim and rest on their back (hence their common name "Backswimmer" or "Water Boatman") and are found under the water surface.[5] Notonecta glauca supports itself under the water surface by using their front legs and mid legs and the back end of its abdomen and rest them on the water surface;[6] They are able to stay under the water surface by water tension, also known as the air-water interface (Surface tension).[7] They use the hind legs as oars; these legs are fringed with hair and, when at rest, are extended laterally like a pair of sculls in a boat.[8] Notonecta glauca will either wait for its prey to pass by or will swim and actively hunt its prey. When the weather is warm, usually in the late summer and autumn, they will fly between ponds.[9][10] Notonecta glauca reproduce in the spring.[2]
| Notonecta glauca | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| Order: | Hemiptera |
| Family: | Notonectidae |
| Genus: | Notonecta |
| Species: | N. glauca |
| Binomial name | |
| Notonecta glauca | |
Eye
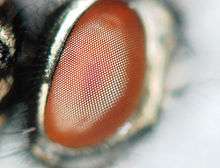
There has been a great deal of research on the eye of N. glauca. These insects use their eyes for both day and night vision, which is used for prey capture and flight when searching for new habitats.[5] Notonecta glauca, like other insects, have a compound eye. Specifically, their eye is an acone-type with corneal structure, which helps them create a sharp image when both in the water and in the air.[11][12][13] The acone is the site of the pupil. Immonen et al. (2014), found that backswimmers are able to see in both day and night light conditions because of:
- their large variations in the peripheral photoreceptor cell properties
- having a roust migration of pigment and photoreceptors
They also found that the green-sensitive peripheral photoreceptors function in a similar way as nocturnal Phasmatodea (or stick insects). To protect their eye from direct sunlight during the day, the pigment cell's diaphragm are condensed, and during the night they open fully to allow as much light in as possible.[5] Notonecta glauca have two photoreceptor subsystems:
- Large and most sensitive peripheral photoreceptors
- Smaller peripheral and central photoreceptors
The first subsystem is sensitive to green light, one of the colours in the visible spectrum. This sensitivity helps the backswimmer see in dimmer light or at night. The second subsystem allows the backswimmer to see in bright light and when in flight.[5] Notonecta glauca pupil (acone) take a different amount of time to adjust to light. It takes the N. glauca approximately 40 minutes for the pupil to adjust to daylight and approximately 50 minutes to adjust to the light at night.[14]
Air retention

Although N. glauca live in the water, they breathe atmosphere air and do not have gills. When these insects are diving or resting under the water surface, they create a film of air that surrounds their body.[15] This air film is also known as a superhydrophobic coating or surface, and it prevents the insect from becoming wet.[4] It also reduces the drag (physics) that is created when diving.[16][17] To be able to create this air film around it, N. glauca is covered in hairy structures, except on its head and legs.[15] There are two types of hairs and air retention is maximized by having both types: setae and microtrichia.[15] The most important part in creating an air film is the density of the hairs.[4] Notonecta glauca has dense microtrichia and their air film can last up to 120 days.[4] The air film cannot last forever because as an insect respires (breathes), the oxygen partial pressure will decrease and nitrogen partial pressure will increase, causing the air bubble to decrease in size.[4]
Model organism
Notonecta glauca is used as a model organism for friction reduction and air retention.[15] Possible applications for this include reduction of drag in ships.
Wave discrimination
Notonecta glauca can discriminate between prey and non-prey, like other backswimmers, by surface waves.[18] Lang (1979), completed an experiment that showed that waves that were created by other backswimmers swimming, emerging, turning and paddling were of a lower frequency (below 40 Hz) compared to waves created by their prey items, who had a frequency between 70–140 Hz. Larval backswimmers were found to create different waves that differed from adult backswimmers, but their frequency were similar to that of adult swimming produced waves (up to 70 Hz).
Water depth
Water depth can affect how N. glauca pick which prey they eat. Males and females both spend a lot of time on the surface of the water where they encounter Culex larvae.
.jpg)
They feed on this prey because there is a decrease in travel cost (having to dive for them) and Culex gives a higher energy rate.[19] Mature females, however, will dive to the bottom of the pond to reach Asellus larvae,

but only in shallow waters.[19] To reach the Asellus larvae there is an increase in the energy used in travel cost (diving) to reach this prey. This behaviour is not consistent with the Optimal foraging theory.[19] It is possible, however, that because mature females are larger than males and immature females, they will have a reduced buoyancy and therefore reduce the energy needed to capture Asellus.[20] Also, mature females have a larger abdominal size, which could support a larger air bubble and allow them to remain submerged for longer.[19] However, if the water depth deepens, mature females will switch and spend more time at the surface and not feed on Asellus, as the deep water will increase the amount of energy needed for diving and staying submerged.[19] The amount of oxygen concentration in the body of water can affect the choice of prey N. glauca pick, as Cockrell (1984), found that when oxygen was at a high dissolved level, N. glauca will spend more time submerged and attacking Asellus.[21]

References
| Wikimedia Commons has media related to Notonecta glauca. |
- Berchi, G.M. (2013). "Checklist and distribution of the family Notonectidae in Romania, with the first record of Notonecta maculata Fabricius, 1794 (Hemiptera: Heteroptera: Nepomorpha)". Zootaxa. 3682 (1): 121–132. doi:10.11646/zootaxa.3682.1.5. PMID 25243278.
- Svensson, B.G; Tailmark, B; Petersson, E (2000). "Habitat Heterogeneity, coexistence and habitat utilization in five backswimmer species (Notonecta spa,; Hemiptera, Notonectidae)". Aquatic Insects. 22 (2): 81–98. doi:10.1076/0165-0424(200004)22:2;1-p;ft081.
- Southwood, Richard; Dennis Leston (1959). Land & Water Bugs of the British Isles. London & New York: Frederick Warne & Co. LTD.
- Balmert, A; Bohn, H.F; Ditsche-Kuru, P; Barhlott, W (2011). "Dry under water: Comparative morphology and functional aspects of air-retaining insect surfaces". Journal of Morphology. 272 (4): 442–451. doi:10.1002/jmor.10921. PMID 21290417.
- Immonen, E.V; Ignatova, I; Gislen, A; Warrant, E; Vahasorinki, M; Weckstöm, M; Frolov, R (2014). "Large variations among photoreceptors as the basis of visual flexibility on the common backswimmer". Proceedings of the Royal Society. 281 (1795): 20141177. doi:10.1098/rspb.2014.1177. PMC 4213611. PMID 25274359.
- Wichard, W; Arens, W; Eisenbeis, G (2002). Biological Atlas of Aquatic Insects. Stenstrup, Denmark: Apollo Books.
- Wachmann, E; Melber, A; Deckert, J (2006). Wanzen. Band 1: Dipsocoromorpha, Nepomorpha, Gerromorpha, Leptopodomorpha. Die Tierwelt Deutschlands.: Keltern: Goecke & Evers. p. 264 p.
-

- Walton, G.A (1935). "Field experiments on the flight of Notonecta maculata Fabr. (Hemipt.)". Trans. Soc. Br. Ent. 2: 137–144.
- Schwind, R (1983). "A polarization-sensitive response of the flying water bug Notonecta glauca to Uv light". J. Comp. Physiol. 150: 87–91. doi:10.1007/bf00605291.
- Fischer, C; Mahner, M; Wachmann, E (2000). "The rhabdom structure in the ommatidia of the Heteroptera (Insecta), and its phylogenetic significance". Zoomorphology. 120: 1–13. doi:10.1007/s004359900018.
- Schwind, R (1980). "Geometrical optics of the Notonecta eye: adaptations to optical environment and way of life". J. Comp. Physiol. 140: 59–68. doi:10.1007/BF00613748.
- Horvath, G (1989). "Geometric optical optimization of the corneal lens of Notonecta glauca". J. Theor. Biol. 139 (3): 389–404. doi:10.1016/S0022-5193(89)80217-6.
- Ro, A.F; Nilsson, D.E (1995). "Pupil adjustments in the eye of the common backswimmer". The Journal of Experimental Biology. 198: 71–77.
- Ditsche-Kuru, P; Schneider, E.S; Melskotte, J.E; Brede, M; Leder, A; Barthlott, W (2011). "Superhydrophopic surfaces of the water bug Notonecta glauca:a model for friction reduction and air retention". Beilstein Journal of Nanotechnology. 2: 137–144. doi:10.3762/bjnano.2.17. PMC 3148060. PMID 21977425.
- McHale, G.; Shirtcliffe, N. J.; Evans, C. R.; Newton, M. I. (2009). "Terminal velocity and drag reduction measurements on superhydrophobic spheres" (PDF). Appl. Phys. Lett. 94 (6): 0641041. doi:10.1063/1.3081420.
- McHale, G.; Newton, M. I.; Shirtcliffe, N. J. (2010). "Immersed superhydrophobic surfaces: Gas exchange, slip and drag reduction properties" (PDF). Soft Matter. 6 (4): 714–719. doi:10.1039/b917861a.
- Lang, H.H (1979). "Surface wave discrimination between prey and non prey by the back swimmer Notonecta gauca L. (Hemiptera, Heteroptera)". Behav. Ecol. Sociobiol. 6 (3): 233–246. doi:10.1007/bf00569205.
- Cockrell, B.J (1984). "Effects of water depth on choice of spatially separated prey by Notonecta glaucaL". Oecologia. 62 (2): 256–261. doi:10.1007/bf00379023. PMID 28310723.
- Alexander, R (1971). Animal mechanics. London, UK: McNeil.
- Cockrell, B.J (1984). "Effect of temperature and oxygenation on predator-prey overlap and prey choice of Notonecta glauca". Journal of Animal Ecology. 53 (2): 519–532. doi:10.2307/4531. JSTOR 4531.