Nigrostriatal pathway
The nigrostriatal pathway is a bilateral dopaminergic pathway in the brain that connects the substantia nigra pars compacta (SNc) in the midbrain with the dorsal striatum (i.e., the caudate nucleus and putamen) in the forebrain. It is one of the four major dopamine pathways in the brain, and is critical in the production of movement as part of a system called the basal ganglia motor loop. Dopaminergic neurons of this pathway release dopamine from axon terminals that synapse onto GABAergic medium spiny neurons (MSNs), also known as spiny projection neurons (SPNs),[1][2] located in the striatum.
| Nigrostriatal pathway | |
|---|---|
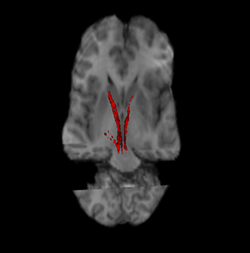 Nigrostriatal pathway (Left and Right in red). | |
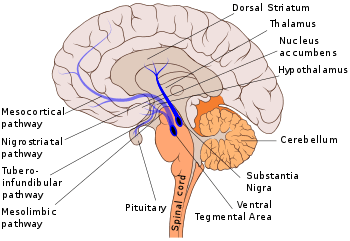 The nigrostriatal pathway is shown here in solid blue, connecting the substantia nigra with the dorsal striatum. | |
| Anatomical terminology |
Degeneration of dopaminergic neurons in the SNc is one of the main pathological features of Parkinson's disease,[3] leading to a marked reduction in dopamine function and the symptomatic motor deficits of Parkinson's disease including hypokinesia, tremors, rigidity, and postural imbalance.
Anatomy
The connection between the substantia nigra pars compacta and the dorsal striatum is mediated via dopaminergic axons.
Substantia nigra pars compacta (SNc)
The substantia nigra is located in the ventral midbrain of each hemisphere. It has two distinct parts, the pars compacta (SNc) and the pars reticulata (SNr). The pars compacta contains dopaminergic neurons from the A9 cell group that forms the nigrostriatal pathway that, by supplying dopamine to the striatum, relays information to the basal ganglia. In contrast, the pars reticulata contains mostly GABAergic neurons.
The SNc is composed of a thin band of cells that overlies the SNr and is situated laterally to the A10 group of dopaminergic neurons in the ventral tegmental area (VTA) that forms the mesolimbic dopamine pathway. The SNc is easily visualized in human brain sections because the dopamine neurons contain a black pigment called neuromelanin which is known to accumulate with age.[4] The dopaminergic cell bodies in the SNc are densely packed with approximately 200,000 to 420,000 dopamine cells in human SNc and 8,000 to 12,000 dopamine cells in mouse SNc.[5] These dopamine cell bodies are localized to one of two chemically defined layers.[6] Those in the upper layer (or dorsal tier) contain a binding protein called calbindin-D28K which can buffer calcium levels inside the cell when it becomes too high or toxic. Dopamine cells in the lower layer (or ventral tier) lacks this protein and are more vulnerable to the effects of neurotoxins (e.g. MPTP) that can cause Parkinson disease-like symptoms.[7][8] The dorsal tier dopamine cells have dendrites that radiate horizontally across the pars compacta, whereas ventral tier dopamine cells have dendrites that extend ventrally into the pars reticulata.[6][9]
Dopaminergic axons
The axons from dopamine neurons emanate from a primary dendrite and project ipsilaterally (on the same side) via the medial forebrain bundle to the dorsal striatum. There is a rough topographical correlation between the anatomical localization of the dopamine cell body within the SNc and the area of termination in the dorsal striatum. Dopaminergic cells in the lateral parts of the SNc project mainly to the lateral and caudal (posterior) parts of the striatum, whereas dopamine cells in the medial SNc project to the medial striatum.[10][9] In addition, dopamine cells in the dorsal tier project to the ventromedial striatum, whereas the ventral tier neurons project to the dorsal caudate nucleus and putamen.[6][9] In general, there is a greater density of dopaminergic input to the dorsolateral striatum.[9]
Each dopamine neuron has an extremely large unmyelinated axonal arborization which can innervate up to 6% of the striatal volume in a rat.[11] Although all SNc dopamine cells project to both the striosome (or patch) and matrix neurochemical compartments of the striatum, most of the axonal territory of a dorsal tier neuron is in the matrix compartment while the majority of the axonal field of ventral tier neurons is in the striosomes.[6][10][11] Nigrostriatal dopamine axons can also give rise to axon collaterals that project to other brain regions. For example, some, SNc nigrostriatal dopamine axons send axon collaterals to the pedunculopontine nucleus, the ventral palladium, subthalamic nucleus, globus pallidus, amygdala and the thalamus.[6][9][12]
A small number of SNc dorsal tier dopamine neurons also project directly to the cortex, although most of the dopaminergic innervation of the cortex comes from the adjacent VTA dopamine neurons.[9]
Dorsal striatum
The dorsal striatum is located in the subcortical region of the forebrain. In primates and other mammals it is divided by the anterior limb of a white matter tract called the internal capsule,[13] into two parts: the caudate nucleus and the putamen.[14] In rodents, the internal capsule is poorly developed such that the caudate and putamen are not separated but form one large entity called the caudate putamen (CPu).[15][16] The majority (about 95%) of cells in the dorsal striatum are GABAergic medium spiny neurons (MSNs) also known as spiny projection neurons (SPNs). Approximately half of these MSNs contain dopamine D1 receptors and project directly to the substantia nigra to form the direct pathway of the basal ganglia, whereas the other half express dopamine D2 receptors that project indirectly to the substantia nigra via the globus pallidus and subthalamic nucleus to form the indirect pathway of the basal ganglia.[17] The remaining 5% of cells are interneurons that are either cholinergic neurons,[18] or one of several types of GABAergic neurons.[19] The axons and dendrites of these interneurons stay within the striatum.
The caudate nucleus and putamen receive excitatory information from all areas of the cerebral cortex.[20] These glutamatergic inputs are generally topographically arranged such that the putamen takes information largely from the sensorimotor cortex whereas the caudate nucleus obtains information largely from the association cortex.[20] In addition, the dorsal striatum receives excitatory inputs from other brain structures like the thalamus,[21] and minor excitatory inputs from the hippocampus and amygdala.
The dorsal striatum contains neurochemically defined compartments called striosomes (also known as patches) that exhibit dense μ-opioid receptor staining embedded within a matrix compartment that contains higher acetylcholinesterase and calbindin-D28K.[22]
The dopaminergic axon terminals of the nigrostriatal pathway synapse onto GABAergic MSNs in the dorsal striatum. They form synapses on the cell body and dendritic shaft regions but mostly on the necks of dendritic spines that also receive glutamatergic input to the heads of the same dendritic spines.[1]
Function
The main function of the nigrostriatal pathway is to influence voluntary movement through basal ganglia motor loops. Along with the mesolimbic and mesocortical dopaminergic pathways the nigrostriatal dopamine pathway can also influence other brain functions including cognition,[23] reward and addiction.[24] Nigrostriatal dopaminergic neurons exhibit tonic and phasic patterns of neuronal firing activity. This can lead to different patterns of dopamine release from the axon terminals in the dorsal striatum and also from the cell body (soma) and dendrites in the SNc and SNr.[25][26] As well as releasing dopamine some axons in the nigrostriatal pathway can also co-release GABA.[27][28]
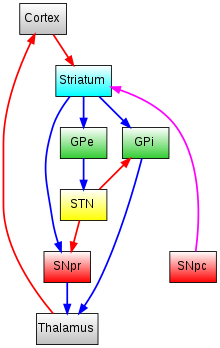
The nigrostriatal pathway influences movement through two pathways, the direct pathway of movement and the indirect pathway of movement.[29][30]
Direct pathway of movement
The direct pathway is involved in facilitation of wanted movements. The projections from dopamine D1 receptor containing medium spiny neurons in the caudate nucleus and putamen synapse onto tonically active GABAergic cells in the substantia nigra pars reticulata and the internal segment of the globus pallidus (GPi) which then project to the thalamus. Because the striatonigral / striatoentopeduncular and nigrothalamic pathways are inhibitory, activation of the direct pathway creates an overall net excitatory on the thalamus and on movement generated by the motor cortex.
Indirect pathway of movement
The indirect pathway is involved in suppressing unwanted movement. The projections from dopamine D2 receptor containing medium spiny neurons in the caudate nucleus and putamen synapse onto tonically active GABAergic cells in the external segment of the globus pallidus (GPe) which then projects to the substantia nigra pars reticulata via the excitatory subthalmic nucleus (STN). Because the striatopallidal and nigrothalamic pathways are inhibitory but the subthalamic to nigra pathway is excitatory, activation of the indirect pathway creates an overall net inhibitory effect on the thalamus and on movement by the motor cortex.
Clinical significance
Parkinson's disease
Parkinson's disease is characterized by severe motor problems, mainly hypokinesia, rigidity, tremors, and postural imbalance.[31] Loss of dopamine neurons in the nigrostriatal pathway is one of the main pathological features of Parkinson's disease.[32] Degeneration of dopamine producing neurons in the substantia nigra pars compacta and the putamen-caudate complex leads to diminished concentrations of dopamine in the nigrostriatal pathway, leading to reduced function and the characteristic symptoms.[33] The symptoms of the disease typically do not show themselves until 80-90% of dopamine function has been lost.
Levodopa-induced dyskinesia
Levodopa-induced dyskinesias (LID) is a complication associated with long-term use of the Parkinson's treatment L-DOPA characterized by involuntary movement and muscle contractions. This disorder occurs in up to 90% of patients after 9 years of treatment. The use of L-DOPA in patients can lead to interruption of nigrostriatal dopamine projections as well as changes in the post-synaptic neurons in the basal ganglia.[34]
Schizophrenia
Presynaptic dopamine metabolism is altered in schizophrenia.[35][36]
Other dopamine pathways
Other major dopamine pathways include:
See also
References
- David Smith, A.; Paul Bolam, J. (1990-07-01). "The neural network of the basal ganglia as revealed by the study of synaptic connections of identified neurones". Trends in Neurosciences. 13 (7): 259–265. doi:10.1016/0166-2236(90)90106-K. ISSN 0166-2236. PMID 1695400.
- Tritsch, NX; Ding, JB; Sabatini, BL (Oct 2012). "Dopaminergic neurons inhibit striatal output through non-canonical release of GABA". Nature. 490 (7419): 262–6. Bibcode:2012Natur.490..262T. doi:10.1038/nature11466. PMC 3944587. PMID 23034651.
- Diaz, Jaime. How Drugs Influence Behavior. Englewood Cliffs: Prentice Hall, 1996.
- Zucca, Fabio A.; Basso, Emy; Cupaioli, Francesca A.; Ferrari, Emanuele; Sulzer, David; Casella, Luigi; Zecca, Luigi (January 2014). "Neuromelanin of the human substantia nigra: an update". Neurotoxicity Research. 25 (1): 13–23. doi:10.1007/s12640-013-9435-y. ISSN 1476-3524. PMID 24155156.
- Brichta, Lars; Greengard, Paul (2014). "Molecular determinants of selective dopaminergic vulnerability in Parkinson's disease: an update". Frontiers in Neuroanatomy. 8: 152. doi:10.3389/fnana.2014.00152. ISSN 1662-5129. PMC 4266033. PMID 25565977.
- Prensa, L.; Giménez-Amaya, J. M.; Parent, A.; Bernácer, J.; Cebrián, C. (2009). "The nigrostriatal pathway: axonal collateralization and compartmental specificity". Journal of Neural Transmission. Supplementum (73): 49–58. doi:10.1007/978-3-211-92660-4_4. ISBN 978-3-211-92659-8. ISSN 0303-6995. PMID 20411767.
- Nemoto, C.; Hida, T.; Arai, R. (1999-10-30). "Calretinin and calbindin-D28k in dopaminergic neurons of the rat midbrain: a triple-labeling immunohistochemical study". Brain Research. 846 (1): 129–136. doi:10.1016/s0006-8993(99)01950-2. ISSN 0006-8993. PMID 10536220.
- Dopeso-Reyes, Iria G.; Rico, Alberto J.; Roda, Elvira; Sierra, Salvador; Pignataro, Diego; Lanz, Maria; Sucunza, Diego; Chang-Azancot, Luis; Lanciego, Jose L. (2014). "Calbindin content and differential vulnerability of midbrain efferent dopaminergic neurons in macaques". Frontiers in Neuroanatomy. 8: 146. doi:10.3389/fnana.2014.00146. ISSN 1662-5129. PMC 4253956. PMID 25520629.
- Haber, S. N. (2014-12-12). "The place of dopamine in the cortico-basal ganglia circuit". Neuroscience. 282: 248–257. doi:10.1016/j.neuroscience.2014.10.008. ISSN 1873-7544. PMC 5484174. PMID 25445194.
- Gerfen, C. R.; Herkenham, M.; Thibault, J. (December 1987). "The neostriatal mosaic: II. Patch- and matrix-directed mesostriatal dopaminergic and non-dopaminergic systems". The Journal of Neuroscience. 7 (12): 3915–3934. doi:10.1523/JNEUROSCI.07-12-03915.1987. ISSN 0270-6474. PMC 6569093. PMID 2891799.
- Matsuda, Wakoto; Furuta, Takahiro; Nakamura, Kouichi C.; Hioki, Hiroyuki; Fujiyama, Fumino; Arai, Ryohachi; Kaneko, Takeshi (2009-01-14). "Single nigrostriatal dopaminergic neurons form widely spread and highly dense axonal arborizations in the neostriatum". The Journal of Neuroscience. 29 (2): 444–453. doi:10.1523/JNEUROSCI.4029-08.2009. ISSN 1529-2401. PMC 6664950. PMID 19144844.
- Prensa, L.; Parent, A. (2001-09-15). "The nigrostriatal pathway in the rat: A single-axon study of the relationship between dorsal and ventral tier nigral neurons and the striosome/matrix striatal compartments". The Journal of Neuroscience. 21 (18): 7247–7260. doi:10.1523/JNEUROSCI.21-18-07247.2001. ISSN 1529-2401. PMC 6762986. PMID 11549735.
- Emos, Marc Christopher; Agarwal, Sanjeev (2019), "Neuroanatomy, Internal Capsule", StatPearls, StatPearls Publishing, PMID 31194338, retrieved 2019-10-06
- Mai, Jürgen K. Atlas of the human brain. Majtanik, Milan,, Paxinos, George, 1944- (4th ed.). Amsterdam. ISBN 9780128028001. OCLC 934406284.
- Coizet, Veronique; Heilbronner, Sarah R.; Carcenac, Carole; Mailly, Philippe; Lehman, Julia F.; Savasta, Marc; David, Oivier; Deniau, Jean-Michel; Groenewegen, Henk J.; Haber, Suzanne N. (March 8, 2017). "Organization of the Anterior Limb of the Internal Capsule in the Rat". The Journal of Neuroscience. 37 (10): 2539–2554. doi:10.1523/JNEUROSCI.3304-16.2017. ISSN 1529-2401. PMC 5354315. PMID 28159909.
- Paxinos, George, 1944- (2013-11-07). The rat brain in stereotaxic coordinates. Watson, Charles, 1943- (Seventh ed.). Amsterdam. ISBN 9780123919496. OCLC 859555862.CS1 maint: multiple names: authors list (link)
- Gerfen, Charles R.; Surmeier, D. James (2011). "Modulation of striatal projection systems by dopamine". Annual Review of Neuroscience. 34: 441–466. doi:10.1146/annurev-neuro-061010-113641. ISSN 1545-4126. PMC 3487690. PMID 21469956.
- Gonzales, Kalynda K.; Smith, Yoland (September 2015). "Cholinergic interneurons in the dorsal and ventral striatum: anatomical and functional considerations in normal and diseased conditions". Annals of the New York Academy of Sciences. 1349 (1): 1–45. Bibcode:2015NYASA1349....1G. doi:10.1111/nyas.12762. ISSN 1749-6632. PMC 4564338. PMID 25876458.
- Tepper, James M.; Koós, Tibor; Ibanez-Sandoval, Osvaldo; Tecuapetla, Fatuel; Faust, Thomas W.; Assous, Maxime (2018). "Heterogeneity and Diversity of Striatal GABAergic Interneurons: Update 2018". Frontiers in Neuroanatomy. 12: 91. doi:10.3389/fnana.2018.00091. ISSN 1662-5129. PMC 6235948. PMID 30467465.
- Haber, Suzanne N. (March 2016). "Corticostriatal circuitry". Dialogues in Clinical Neuroscience. 18 (1): 7–21. ISSN 1958-5969. PMC 4826773. PMID 27069376.
- Smith, Yoland; Galvan, Adriana; Ellender, Tommas J.; Doig, Natalie; Villalba, Rosa M.; Huerta-Ocampo, Icnelia; Wichmann, Thomas; Bolam, J. Paul (2014). "The thalamostriatal system in normal and diseased states". Frontiers in Systems Neuroscience. 8: 5. doi:10.3389/fnsys.2014.00005. ISSN 1662-5137. PMC 3906602. PMID 24523677.
- Brimblecombe, Katherine R.; Cragg, Stephanie J. (February 15, 2017). "The Striosome and Matrix Compartments of the Striatum: A Path through the Labyrinth from Neurochemistry toward Function". ACS Chemical Neuroscience. 8 (2): 235–242. doi:10.1021/acschemneuro.6b00333. ISSN 1948-7193. PMID 27977131.
- Boot, Nathalie; Baas, Matthijs; van Gaal, Simon; Cools, Roshan; De Dreu, Carsten K. W. (July 2017). "Creative cognition and dopaminergic modulation of fronto-striatal networks: Integrative review and research agenda". Neuroscience and Biobehavioral Reviews. 78: 13–23. doi:10.1016/j.neubiorev.2017.04.007. ISSN 1873-7528. PMID 28419830.
- Wise, RA (October 2009). "Roles for nigrostriatal--not just mesocorticolimbic--dopamine in reward and addiction". Trends in Neurosciences. 32 (10): 517–524. doi:10.1016/j.tins.2009.06.004. PMC 2755633. PMID 19758714.
- Rice, M. E.; Patel, J. C.; Cragg, S. J. (2011-12-15). "Dopamine release in the basal ganglia". Neuroscience. 198: 112–137. doi:10.1016/j.neuroscience.2011.08.066. ISSN 1873-7544. PMC 3357127. PMID 21939738.
- Rice, Margaret E.; Patel, Jyoti C. (2015-07-05). "Somatodendritic dopamine release: recent mechanistic insights". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1672): 20140185. doi:10.1098/rstb.2014.0185. ISSN 1471-2970. PMC 4455754. PMID 26009764.
- Tritsch, Nicolas X.; Granger, Adam J.; Sabatini, Bernardo L. (March 2016). "Mechanisms and functions of GABA co-release". Nature Reviews. Neuroscience. 17 (3): 139–145. doi:10.1038/nrn.2015.21. ISSN 1471-0048. PMC 6980171. PMID 26865019.
- Trudeau, Louis-Eric; Hnasko, Thomas S.; Wallén-Mackenzie, Asa; Morales, Marisela; Rayport, Steven; Sulzer, David (2014). "The multilingual nature of dopamine neurons". Progress in Brain Research. 211: 141–164. doi:10.1016/B978-0-444-63425-2.00006-4. ISBN 9780444634252. ISSN 1875-7855. PMC 4565795. PMID 24968779.
- Kravitz, Alexxai V.; Kreitzer, Anatol C. (June 2012). "Striatal mechanisms underlying movement, reinforcement, and punishment". Physiology. 27 (3): 167–177. doi:10.1152/physiol.00004.2012. ISSN 1548-9221. PMC 3880226. PMID 22689792.
- Kravitz, Alexxai V.; Freeze, Benjamin S.; Parker, Philip R. L.; Kay, Kenneth; Thwin, Myo T.; Deisseroth, Karl; Kreitzer, Anatol C. (2010-07-29). "Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry". Nature. 466 (7306): 622–626. Bibcode:2010Natur.466..622K. doi:10.1038/nature09159. ISSN 1476-4687. PMC 3552484. PMID 20613723.
- Cenci, Angela M (2006). "Post- versus presynaptic plastic ity in L-DOPA-induced dyskinesia" (PDF). Journal of Neurochemistry. 99 (2): 381–92. doi:10.1111/j.1471-4159.2006.04124.x. PMID 16942598.
- Deumens, Ronald (21 June 2002). "Modeling Parkinson's Disease in Rats: An Evaluation of 6-OHDA Lesions of the Nigrostriatal Pathway". Experimental Neurology. 175 (2): 303–17. doi:10.1006/exnr.2002.7891. PMID 12061862.
- Groger, Adraine (8 January 2014). "Dopamine Reduction in the Substantia Nigra of Parkinson's Disease Patients Confirmed by In Vivo Magnetic Resonance Spectroscopic Imaging". PLoS ONE. 9 (1): e84081. Bibcode:2014PLoSO...984081G. doi:10.1371/journal.pone.0084081. PMC 3885536. PMID 24416192.
- Niethammer, Martin (May 2012). "Functional Neuroimaging in Parkinson's Disease". Cold Spring Harbor Perspectives in Medicine. 2 (5): a009274. doi:10.1101/cshperspect.a009274. PMC 3331691. PMID 22553499.
- Fusar-Poli, Paolo; Meyer-Lindenberg, Andreas (1 January 2013). "Striatal presynaptic dopamine in schizophrenia, part II: meta-analysis of [(18)F/(11)C]-DOPA PET studies". Schizophrenia Bulletin. 39 (1): 33–42. doi:10.1093/schbul/sbr180. ISSN 1745-1701. PMC 3523905. PMID 22282454.
- Weinstein, Jodi J.; Chohan, Muhammad O.; Slifstein, Mark; Kegeles, Lawrence S.; Moore, Holly; Abi-Dargham, Anissa (1 January 2017). "Pathway-Specific Dopamine Abnormalities in Schizophrenia". Biological Psychiatry. 81 (1): 31–42. doi:10.1016/j.biopsych.2016.03.2104. ISSN 1873-2402. PMC 5177794. PMID 27206569.