Karrikin
Karrikins are a group of plant growth regulators found in the smoke of burning plant material.[1][2] Karrikins help stimulate seed germination and plant development because they mimic a signaling hormone known as strigolactone. Strigolactones are hormones that help increase growth of symbiotic arbuscular mycorrhizal fungi in the soil, which enhances plant growth and leads to an increase in plant branching. [3][4]
Smoke from wildfires or bushfires has been known for a long time to stimulate the germination of seeds.[5][6] In 2004, the butenolide karrikinolide (KAR1) was shown to be responsible for this effect.[7] Later, several closely related compounds were discovered in smoke, and are collectively known as karrikins.[2]
Chemical synthesis
Karrikins are formed by the heating or combustion of carbohydrates, including sugars and polysaccharides, mainly cellulose.[8] When plant material burns, these carbohydrates convert to karrikins. The pyran moiety of karrikins is probably directly derived from a pyranose sugar. There is no evidence that karrikins occur naturally in plants, but it has been postulated that karrikin-like molecules do.[9]
Karrikin taxonomy
It has long been known that compounds released from smoke stimulate seed germination. To identify the active compounds that contribute to seed germination activity, smoke compounds were separated by liquid fractionation and were each tested for their effects on seed germination activity. Bioassays identified several related compounds that were named karrikins.[10]
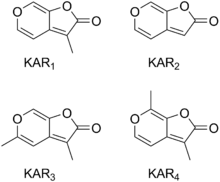
Six karrikins have so far been discovered in smoke, and they are designated KAR1, KAR2, KAR3, KAR4, KAR5 and KAR6. KAR1 to KAR4 are the most active karrikins.[11] KAR1 is also known as karrikinolode and was the first karrikin to be discovered.[10]
Mode of action
Karrikins are released into the air upon the burning of plants. Subsequently, karrikins then get deposited on the soil surface and stimulate seed germination after rainfall. Since karrikins are released from smoke, they are released in huge quantities. [12] Interestingly, some plants which are known as "fire-followers" are unable to germinate without karrikins. Fire-followers need rain after massive fires in order to germinate; this means that they may remain dormant and viable for decades until the right combination of fire occur in proper succession.[10]
Etymology
The first karrikin discovered, abbreviated as KAR1, was initially named gavinone in reference to its discovery by chemist Gavin Flematti. After consulting with an etymologist, Flematti proposed changing the name of the molecule and its related compounds to karrikin. One of the first recorded Western Australian Noongar words for 'smoke' from the Perth area in the 1830s, is 'karrik' [2][13][14][15] .
The response to karrikins
Karrikins produced by bushfires occur largely in the ash at the site of the fire. Rains occurring after the fire wash the karrikins into the soil where dormant seeds reside. The karrikins and water can provide a 'wake-up call’ for such seeds, triggering germination of the soil seed bank. The plants that depend on karrikins to grow are known as "fire-followers" [16], they emerge grow quickly, flower and produce new seeds, which fall to the ground. These seeds can remain in the soil for decades, until the next fire produces fresh karrikins. Plants with this lifestyle are known as fire ephemerals. They thrive because the fire removes competing vegetation and provides nutrients and light for the emerging seedlings. Plants in many families respond to smoke and karrikins, suggesting that this response has evolved independently in different groups.[11]
Fire-followers are not the only plants that respond to karrikins. Seeds from a number of different flowering families like tomatoes, lettuce, and trees respond to karrikin signaling. [17]. Interestingly, other studies have found that seed of ostensibly fire-adapted species do not display a sensitivity to karrikins. [18] The difference between fire-followers and plants that respond to karrikins is their dependence on karrikins. [17] Plants' response to karrikins is fundamental because karrikins mimic the strigolactone hormones which are originally required for growth in plants. Fire-followers, on the other hand, have fine-tuned their responses according to the availability of karrikins. [17]
Structure and physicochemical properties
Carbon, hydrogen, and oxygen make up the two ring structures found in karrikins, one of which is a six-membered, heterocyclic ring with a molecular formula of C5H6O known as pyran, and the other is a five-membered lactone ring known as a butenolide.[19]
Karrikins easily dissolve in water, they are transparent, and have a melting point of 118–119 °C. [19] However, they are unstable at very high temperatures and during common daylight, which means that they decay more rapidly than common active compounds which are not sensitive to sunlight. [20][19]
Mechanism of action
The mode of action of karrikins has been largely determined using the genetic resources of Arabidopsis thaliana. Perception of karrikins by Arabidopsis requires an alpha/beta-fold hydrolase named KARRIKIN-INSENSITIVE-2 (KAI2).[21] The KAI2 protein has a catalytic triad of amino acids which is essential for activity, consistent with the hypothesis that KAI2 hydrolyses its ligand.[22][23] This model is consistent with the perception of the chemically related strigolactone hormones which involves hydrolysis by their receptor protein DWARF14, an alpha/beta hydrolase related to KAI2.[21][24] The question of whether karrikins act directly in plants is controversial. While some studies suggest that karrikins can bind directly to KAI2 protein,[25] others do not support this.[23] It is possible that karrikins produced by wildfires are converted to a different compound by the plant, before interaction with KAI2. The ability of different plants to carry out this conversion could partly explain differences in their ability to respond to karrikins and to smoke.
Signalling
The activity of karrikins requires an F-box protein named MORE AXILLARY GROWTH-2 (MAX2) in Arabidopsis.[26] This protein is also required for strigolactone signaling in Arabidopsis. Homologs of MAX2 are also required for strigolactone signaling in rice (known as DWARF3) petunia (DAD2) and pea (RMS4). Karrikin signaling also requires a protein named SUPPRESSOR OF MORE AXILARY GROWTH2-1 (SMAX1) [27] which is a homolog of the DWARF53 protein required for strigolactone signaling in rice.[28][29] SMAX1 and DWARF53 proteins could be involved in the control of cellular functions such as transport or transcription.[24] The present model for karrikin and strigolactone signaling involves interaction of KAI2 or DWARF14 with SMAX1 or DWARF53 proteins respectively, which targets those proteins for ubiquitination and destruction.[30]
Studies have shown that Arabidopsis responds to the two signals; KAR1, and KAR2. The two genes, MORE AXILLARY GROWTH2 (MAX2) and KARRIKIN-INSENSITIVE2 (KAI2) are essential for understanding the actions of karrikins and were discovered in Arabidopsis mutants which failed to respond to karrikins. In rice, strigolactones interact with the F-box protein knowns as DWARF3 upon their hydrolysis by the DWARF14 (also known as D14-type proteins). This interaction targets the ubiquitination and destruction of proteins which are responsible for different aspect of plant growth, like the outgrowth of lateral shoots. This means that strigolactones, upon their interaction with D3 and D14; ubiquinate, and destroy proteins like DWARF53, which are responsible for the outgrowth of lateral shoots, and for the inhibition of stem thickening and root branching.[31] In Arabidopsis, Karrikins work in a similar way to strigolactones; they require homologous proteins known as KARRIKIN-INSENSITIVE1 (KAI1 or MAX2) in order to be able to interact with KARRIKIN-INSENSITIVE2 which is responsible for hypocotyl elongation and the inhibition of seed germination. The ubiquination of KAI2, therefore stimulate seed germination and inhibits hypocotyl elongation.[32][33] Karrikins could be used as agricultures, considering the environmental challenges that are occurring nowadays.[34]
Effects on plant growth
Karrikins not only stimulate seed germination, but are reported to increase seedling vigour.[35] In Arabidopsis, karrikins influence seedling photomorphogenesis, resulting in shorter hypocotyls and larger cotyledons. Such responses could provide seedlings with an advantage as they emerge into the post-fire landscape. The KAI2 protein is also required for leaf development, implying that karrikins could influence other aspects of plant growth.
Evolution
The gene for KAI2 protein is present in lower plants including algae and mosses, whereas the DWARF14 protein evolved with seed plants, probably as a result of duplication of KAI2 followed by functional specialisation. Karrikin signaling could have evolved with seed plants as a result of the divergence of KAI2 and DWARF14 functions, possibly during the Cretaceous period when fires were common on Earth.[36]
Response to wildfires
Karrikins are produced by wildfires but all seed plants contain KAI2 proteins, raising the question of the usual function of this protein. There is compelling evidence that plants contain an endogenous compound that is perceived by KAI2 to control seed germination and plant development, but this compound is neither a karrikin nor a strigolactone.[23]
References
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13 (1): 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- Chiwocha SD, Dixon KW, Flematti GR, Ghisalberti EL, Merritt DJ, Nelson DC, et al. (2009-10-01). "Karrikins: A new family of plant growth regulators in smoke". Plant Science. 177 (4): 252–256. doi:10.1016/j.plantsci.2009.06.007.
- "Strigolactone - Latest research and news | Nature". www.nature.com. Retrieved 2020-04-26.
- Andreo-Jimenez B, Ruyter-Spira C, Bouwmeester HJ, Lopez-Raez JA (2015-09-01). "Ecological relevance of strigolactones in nutrient uptake and other abiotic stresses, and in plant-microbe interactions below-ground". Plant and Soil. 394 (1): 1–19. doi:10.1007/s11104-015-2544-z.
- Halford B (2010). "Smoke Signals". Chemical & Engineering News. 88 (15): 37–8. doi:10.1021/cen-v088n015.p037.
- Nelson DC, Flematti GR, Ghisalberti EL, Dixon KW, Smith SM (2012-06-02). "Regulation of seed germination and seedling growth by chemical signals from burning vegetation". Annual Review of Plant Biology. 63 (1): 107–30. doi:10.1146/annurev-arplant-042811-105545. PMID 22404467.
- Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (August 2004). "A compound from smoke that promotes seed germination". Science. 305 (5686): 977. doi:10.1126/science.1099944. PMID 15247439.
- Flematti GR, Scaffidi A, Dixon KW, Smith SM, Ghisalberti EL (February 2011). "Production of the seed germination stimulant karrikinolide from combustion of simple carbohydrates". Journal of Agricultural and Food Chemistry. 59 (4): 1195–8. doi:10.1021/jf1041728. PMID 21280622.
- Gutjahr C, Gobbato E, Choi J, Riemann M, Johnston MG, Summers W, et al. (December 2015). "Rice perception of symbiotic arbuscular mycorrhizal fungi requires the karrikin receptor complex". Science. 350 (6267): 1521–4. Bibcode:2015Sci...350.1521G. doi:10.1126/science.aac9715. PMID 26680197.
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13: 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- Nelson DC, Flematti GR, Ghisalberti EL, Dixon KW, Smith SM (2012-01-01). "Regulation of seed germination and seedling growth by chemical signals from burning vegetation". Annual Review of Plant Biology. 63 (1): 107–30. doi:10.1146/annurev-arplant-042811-105545. PMID 22404467.
- Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (August 2004). "A compound from smoke that promotes seed germination". Science. 305 (5686): 977. doi:10.1126/science.1099944. PMID 15247439.
- Chemistry : human activity, chemical reactivity. [Place of publication not identified]: Brooks Cole. 2014. ISBN 978-1305284203. OCLC 920019241.
- Flematti GR, Ghisalberti EL, Dixon KW, Trengove RD (October 2009). "Identification of alkyl substituted 2H-furo[2,3-c]pyran-2-ones as germination stimulants present in smoke". Journal of Agricultural and Food Chemistry. 57 (20): 9475–80. doi:10.1021/jf9028128. PMID 19785418.
- Nelson DC, Riseborough JA, Flematti GR, Stevens J, Ghisalberti EL, Dixon KW, Smith SM (February 2009). "Karrikins discovered in smoke trigger Arabidopsis seed germination by a mechanism requiring gibberellic acid synthesis and light". Plant Physiology. 149 (2): 863–73. doi:10.1104/pp.108.131516. PMC 2633839. PMID 19074625.
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13: 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13: 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- George N (August 2009). "Does karrikinolide improve the germination and seedling vigour of switchgrass?". Seed Science and Technology. 37 (1): 251–254. doi:10.15258/sst.2009.37.1.30. PMID 15247439.
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13: 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- Scaffidi A, Waters MT, Skelton BW, Bond CS, Sobolev AN, Bythell-Douglas R, et al. (May 2012). "Solar irradiation of the seed germination stimulant karrikinolide produces two novel head-to-head cage dimers". Organic & Biomolecular Chemistry. 10 (20): 4069–73. doi:10.1039/c2ob25090j. PMID 22514031.
- Waters MT, Nelson DC, Scaffidi A, Flematti GR, Sun YK, Dixon KW, Smith SM (April 2012). "Specialisation within the DWARF14 protein family confers distinct responses to karrikins and strigolactones in Arabidopsis". Development. 139 (7): 1285–95. doi:10.1242/dev.074567. PMID 22357928.
- Waters MT, Scaffidi A, Flematti G, Smith SM (May 2015). "Substrate-Induced Degradation of the α/β-Fold Hydrolase KARRIKIN INSENSITIVE2 Requires a Functional Catalytic Triad but Is Independent of MAX2". Molecular Plant. 8 (5): 814–7. doi:10.1016/j.molp.2014.12.020. PMID 25698586.
- Waters MT, Scaffidi A, Moulin SL, Sun YK, Flematti GR, Smith SM (July 2015). "A Selaginella moellendorffii Ortholog of KARRIKIN INSENSITIVE2 Functions in Arabidopsis Development but Cannot Mediate Responses to Karrikins or Strigolactones". The Plant Cell. 27 (7): 1925–44. doi:10.1105/tpc.15.00146. PMC 4531350. PMID 26175507.
- Smith SM, Li J (October 2014). "Signalling and responses to strigolactones and karrikins". Current Opinion in Plant Biology. SI: Cell signalling and gene regulation. 21: 23–29. doi:10.1016/j.pbi.2014.06.003. PMID 24996032.
- Guo Y, Zheng Z, La Clair JJ, Chory J, Noel JP (May 2013). "Smoke-derived karrikin perception by the α/β-hydrolase KAI2 from Arabidopsis". Proceedings of the National Academy of Sciences of the United States of America. 110 (20): 8284–9. Bibcode:2013PNAS..110.8284G. doi:10.1073/pnas.1306265110. PMC 3657771. PMID 23613584.
- Nelson DC, Scaffidi A, Dun EA, Waters MT, Flematti GR, Dixon KW, et al. (May 2011). "F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana". Proceedings of the National Academy of Sciences of the United States of America. 108 (21): 8897–902. Bibcode:2011PNAS..108.8897N. doi:10.1073/pnas.1100987108. PMC 3102411. PMID 21555559.
- Stanga JP, Smith SM, Briggs WR, Nelson DC (September 2013). "SUPPRESSOR OF MORE AXILLARY GROWTH2 1 controls seed germination and seedling development in Arabidopsis". Plant Physiology. 163 (1): 318–30. doi:10.1104/pp.113.221259. PMC 3762653. PMID 23893171.
- Jiang L, Liu X, Xiong G, Liu H, Chen F, Wang L, et al. (December 2013). "DWARF 53 acts as a repressor of strigolactone signalling in rice". Nature. 504 (7480): 401–5. Bibcode:2013Natur.504..401J. doi:10.1038/nature12870. PMC 5802366. PMID 24336200.
- Zhou F, Lin Q, Zhu L, Ren Y, Zhou K, Shabek N, et al. (December 2013). "D14-SCF(D3)-dependent degradation of D53 regulates strigolactone signalling". Nature. 504 (7480): 406–10. Bibcode:2013Natur.504..406Z. doi:10.1038/nature12878. PMC 4096652. PMID 24336215.
- Bennett T, Leyser O (December 2014). "Strigolactone signalling: standing on the shoulders of DWARFs". Current Opinion in Plant Biology. SI: Cell biology. 22: 7–13. doi:10.1016/j.pbi.2014.08.001. PMID 25179782.
- Smith SM (March 2014). "Q&A: What are strigolactones and why are they important to plants and soil microbes?". BMC Biology. 12 (1): 19. doi:10.1186/1741-7007-12-19. PMC 3994223. PMID 24685292.
- Flematti GR, Dixon KW, Smith SM (December 2015). "What are karrikins and how were they 'discovered' by plants?". BMC Biology. 13: 108. doi:10.1186/s12915-015-0219-0. PMC 4687367. PMID 26689715.
- Meng Y, Shuai H, Luo X, Chen F, Zhou W, Yang W, Shu K (2017-01-24). "Karrikins: Regulators Involved in Phytohormone Signaling Networks during Seed Germination and Seedling Development". Frontiers in Plant Science. 7: 2021. doi:10.3389/fpls.2016.02021. PMC 5258710. PMID 28174573.
- Antala M, Sytar O, Rastogi A, Brestic M (December 2019). "Potential of Karrikins as Novel Plant Growth Regulators in Agriculture". Plants. 9 (1): 43. doi:10.3390/plants9010043. PMC 7020145. PMID 31888087.
- van Staden J, Sparg SG, Kulkarni MG, Light ME (2006-08-01). "Post-germination effects of the smoke-derived compound 3-methyl-2H-furo[2,3-c]pyran-2-one, and its potential as a preconditioning agent". Field Crops Research. 98 (2–3): 98–105. doi:10.1016/j.fcr.2005.12.007.
- He T, Pausas JG, Belcher CM, Schwilk DW, Lamont BB (May 2012). "Fire-adapted traits of Pinus arose in the fiery Cretaceous". The New Phytologist. 194 (3): 751–9. doi:10.1111/j.1469-8137.2012.04079.x. hdl:10261/48120. PMID 22348443.