Hematopoietic stem cell niche
Many human blood cells, such as red blood cells (RBCs), immune cells, and even platelets all originate from the same progenitor cell, the hematopoietic stem cell (HSC). As these cells are short-lived, there needs to be a steady turnover of new blood cells and the maintenance of an HSC pool. This is broadly termed hematopoiesis.[1] This event requires a special environment, termed the hematopoietic stem cell niche, which provides the protection and signals necessary to carry out the differentiation of cells from HSC progenitors.[1] This niche relocates from the yolk sac to eventually rest in the bone marrow of mammals. Many pathological states can arise from disturbances in this niche environment, highlighting its importance in maintaining hematopoiesis.[1]
Hematopoiesis
Hematopoiesis involves a series of differentiation steps from one progenitor cell to a more committed cell type, forming the recognizable tree seen in the adjacent diagram. Pluripotent long-term (LT)-HSCs self-renew to maintain the HSC pool, as well as differentiate into short-term (ST)-HSCs.[1] Through various knock-out models, several transcription factors have been found to be essential in this differentiation, such as RUNX1 and TAL1 (also known as SCL).[2][3]
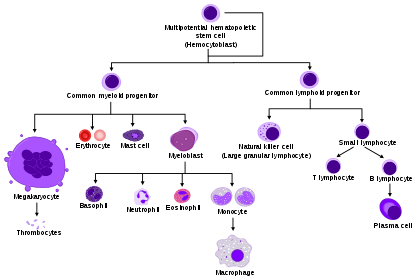
ST-HSCs can then differentiate into either the common myeloid progenitor (CMP) or the common lymphoid progenitor (CLP). The CLP then goes on to differentiate into more committed lymphoid precursor cells. The CMP can then further differentiate into the megakaryocyte–erythroid progenitor cell (MEP), which goes on to make RBCs and platelets, or the granulocyte/macrophage progenitor (GMP), which gives rise to the granulocytes of the innate immune response. MEP differentiation was found to be contingent upon the transcription factor GATA1, whereas GMP differentiation needs SPI1. When expression of either was inhibited by morpholino in zebrafish, the other lineage programming pathway resulted.[4][5]
There are 2 types of hematopoiesis that occur in humans:
- Primitive hematopoiesis – blood stem cells differentiate into only a few specialized blood lineages (typically isolated to early fetal development).
- Definitive hematopoiesis – multipotent HSCs appear (occurs through the majority of human lifetime).
Historical development of the theory
The pioneering work of Till and McCulloch in 1961 experimentally confirmed the development of blood cells from a single precursor hematopoietic stem cell (HSC), creating the framework for the field of hematopoiesis to be studied over the following decades.[6] In 1978, after observing that the prototypical colony-forming stem cells were less capable at replacing differentiated cells than bone marrow cells injected into irradiated animals, Schofield proposed that a specialized environment in the bone marrow allows these precursor cells to maintain their cellular reconstitution potential.[7]
During this time, the field exploded with studies aimed at determining the components of the "hematopoietic stem cell niche" that made this possible. Dexter observed that mesenchymal stromal cells could maintain early HSCs ex vivo, and both Lord and Gong showed that these cells localized to the endosteal margins in long bones.[8][9][10] These studies and others supported the idea that bone cells create the HSC niche, and all the research that elucidated this specialized hematopoietic microenvironment stemmed from these landmark studies.
Niche localization through early fetal development
Yolk sac and the hemangioblast theory
Despite the vast work done in this field, there is still controversy over the origins of definitive HSCs. Primitive hematopoiesis is first found in the blood islands (Pander's islands) of the yolk sac at E7.5 (embryonic day 7.5) in mice and 30dpc (30 days post-conception) in humans. As the embryo requires rapid oxygenation due to its high mitotic activity, these islands are the main source of red blood cell (RBC) production via fusing endothelial cells (ECs) with the developing embryonic circulation.
The hemangioblast theory, which posits that the RBCs and ECs derive from a common progenitor cell, was developed as researchers observed that receptor knockout mice, such as Flk1-/-, exhibited defective RBC formation and vessel growth.[11] A year later, Choi showed that blast cells derived from embryonic stem (ES) cells displayed common gene expression of both hematopoietic and endothelial precursors.[12] However, Ueno and Weissman provided the earliest contradiction to the hemangioblast theory when they saw that distinct ES cells mixed into a blastocyst resulted in more than 1 ES cell contributing to the majority of the blood islands found in the resultant embryo.[13] Other studies done in zebrafish have more soundly indicated the existence of the hemangioblast.[14][15][16] While the hemangioblast theory appears to be generally supported, most of the studies done have been in vitro, indicating a need for in vivo studies to elucidate its existence.[17]
Aorta-gonad-mesonephros region
Definitive hematopoiesis then occurs later in the aorta-gonad-mesonephros (AGM), a region of embryonic mesoderm that develops into the ventral wall of the dorsal aorta, at E10.5 in mice and 4wpc (4 weeks post-conception) in humans.[18] New HSCs either enter the aortic circulation or remain within the endothelium. While Notch 1 has been found to stimulate aortic HSC production, Runx1 overexpression in the zebrafish mutant mindbomb that lacks Notch signaling rescues HSC production, suggesting Runx1 is downstream of Notch1.[19][20] Hedgehog signaling is also required for HSC production in the AGM.[21] ECs located in this niche have been found to support new HSCs through the upregulation of factors such as p57 and IGF2.[22] The relocalization of hemogenic endothelium coincides with the migration of distinct endothelial precursors to the AGM.[23]
Niche relocation through late fetal development
Placenta and the fetal liver
Hematopoiesis then moves from the AGM to the placenta and fetal liver at E11.5 in mice and 5wpc in humans. While the engraftment of HSCs at these sites are still being elucidated, the interaction between the chemokine CXCL12 expressed by stromal cells and its receptor CXCR4 expressed on HSCs has been proposed as one mechanism.[24][25] In addition, the cytokine-receptor binding of SCF and KIT have been recognized for its importance in HSC function and amplification of the chemotactic induction of CXCL12.[26][27]
Additional factors that are important in HSC migration during this period are Integrins, N-cadherin, and Osteopontin that can stimulate Wnt signaling in HSCs.[28][29] Transcription factors such as PITX2 must be expressed in stromal cells to support normal HSC function.[30] Like with the AGM, the relocation of fetal liver HSCs coincides with the differentiation of functional units, in this case hepatoblasts to hepatocytes.[31] Mice have also shown hematopoietic activity in the umbilical arteries and the allantois, in which HSCs and endothelial cells are colocalized.[32]
Bone marrow
Hematopoiesis then moves to the bone marrow at E18 in mice and 12wpc in humans, where it will reside permanently for the remainder of the individual's lifetime. In mice, there is a shift from the fetal liver to the spleen at E14, where it persists for many weeks postnatally while it occurs simultaneously in the bone marrow.[33] This relocalization is thought to be supported by the development of osteoblast and chondrocyte precursor cells capable of forming an HSC niche.[34] In addition to the previously mentioned signals that induce HSC migration, TIE2-angiopoietin and CD44-E-cadherin binding appears to be important for this event to occur, as well as for the retention of these HSCs once they are in the bone marrow.[35][36]
HSCs in the bone marrow do not show the same characteristics as those in other niches. HSCs in fetal liver display increased cell divisions, whereas adult bone marrow HSCs are mostly quiescent.[17] This difference stems, in part, from signaling discrepancies in the two niches. Sox17 has been identified as crucial for the generation of fetal, but not adult, HSCs.[37] Inactivation of Runx1 in adult HSCs does not impair function, but rather prevents the differentiation of specific lineages.[38] Differences such as these in the reactivity of HSCs from different niches suggests that the signaling found there is not the same.
Composition of the bone marrow niche
In addition to the cytokines and cell signaling molecules mentioned above, the HSC niche in the bone marrow provides soluble factors, forces, and cell-mediated interactions necessary to maintain the hematopoietic potential of the stem cells located there. This niche is generally separated into 2 sections:
- Endosteal niche-the outer edge of the bone marrow that contains osteocytes, bone matrix, and quiescent HSCs.
- Perivascular niche-the inner core of the bone marrow that contains actively dividing HSCs, sinusoidal endothelium, CARs (CXCL12-abundant reticular cells), and MSCs (Mesenchymal stem cells).
Acellular factors
Recent studies have used hypoxic-staining dyes, such as Hoechst stain, to show that quiescent LT-HSCs and osteoblasts are found in hypoxic and poorly perfused areas of the bone marrow, while ECs and MSCs were found in well-perfused areas.[39][40] However, this hypoxia may be only caused in part by the niche environment, and the HSCs themselves may be maintaining their hypoxic environment in order to remain quiescent.[41] This oxygen tension upregulates HIF1A, which shifts energy production to glycolysis, allowing for the cell to survive in oxygen-poor surroundings.[42] Indeed, deletion of HIF1A increases HSC proliferation and eventually depletes the LT-HSC storage pool.[43] This suggests that the hypoxic environment of the bone marrow, partially determined by the distance from the sinusoids of the perivascular niche, maintains the quiescent state of LT-HSCs in an effort to retain stem cells with differentiation potential.
It has also been found that calcium ions can act as chemotactic signals to HSCs via the G protein–coupled receptor (GPCR) calcium-sensing receptor (CaSR). CaSR knockout mice have shown hematopoietic cells in the circulation and spleen, but few in the bone marrow, indicating this receptor's importance in this particular niche.[44] Conversely, stimulation of HSC CaSR via its agonist cinacalcet increases migration and engraftment of these cells in the bone marrow.[45] Finally, osteoclast inhibition by the bisphosphonate alendronate has correlated with decreased HSCs and bone marrow engraftment.[46] Taken together, these results suggest that the high calcium ion concentration found in the endosteal niche due to osteoclast activity acts as a homing signal to HSCs for engraftment into the bone marrow via the CaSR.
Thirdly, the shearing forces experienced by HSCs from circulating cells has been suggested to play a role in hematopoietic activation. Fetal HSCs in the AGM have shown upregulated Runx1 in response to these forces, which would result in significant hematopoietic regulation in these cells.[47] Despite the differences between the AGM and bone marrow, both are subjected to the circulation, and it is entirely possible that these same forces exist in this adult stem cell niche. Other characteristics, such as strain, geometry, and ligand profiles of the extracellular matrix (ECM) have been suggested as important in the maintenance of stem cell potential in these niches.[48] Finally, the elasticity modulus of the ECM, partially provided by MSCs in the bone marrow, has been shown to direct the differentiation and activity of stem cells nearby.[49][50] The landscape of the HSC niche in the bone marrow is constantly changing, and the acellular factors, as much as the cellular factors, are beginning to reveal the complexity of hematopoietic regulation.
Cellular factors
Osteoblasts, bone-forming cells, interact with HSCs and provide proliferative signals. Studies that have increased or decreased osteoblasts have shown a similar increase or decrease, respectively, in the number of HSCs.[51][52] Coculturing endosteal cells with HSCs was also found to be sufficient in maintaining their differentiation potential long-term, presumably through the secretion of the cell signaling molecules previously mentioned.[53][54] These HSCs that interact with the endosteal osteoblasts display a quiescent phenotype, as shown in both ex vivo and in vivo imaging studies, whereas HSCs that are more actively dividing show less interaction.[55][56][57] These results of less active HSCs interacting with the endosteal niche agree with previous results looking at the activity state of HSCs throughout the bone marrow.
In addition to osteoblasts, HSCs interact with many mesenchymal cells as they make their way to the sinusoids in the perivascular niche. Removal of nestin-expressing MSCs has shown a significant decrease in LT-HSCs.[58] These cells secrete high levels of CXCL12 and closely associate with sympathetic nerves that influence cytokine-induced migration of HSCs.[59][60] Similar to these cells, CAR cells have correlated with reduced HSCs and LT-HSC activity when ablated.[61] One difference between these cell types, despite the similarity in function, is that CAR cells may be found in both endosteal and perivascular niches, while nestin-positive MSCs are found exclusively in the perivascular niche.
Finally, both ECs and adipocytes have been suggested to influence HSC activity in the bone marrow. Studies using antibody-mediated disruption of the VEGF receptors on ECs correlated with poor engraftment of donor cells.[62] HUVECs, or endothelial cells isolated from umbilical veins, that were programmed via viral gene manipulation to signal through the Notch and Angiopoietin pathways were found to support and maintain LT-HSCs.[63] Despite the difficulty in isolating sinusoidal ECs, experimental evidence suggests that these cells may play a role in regulating HSCs. Current research suggests that adipocytes in the bone marrow negatively regulate HSC activity. HSCs isolated from adipocyte-rich vertebrae displayed decreased activity.[64] In addition, isolation of adipocyte-containing stromal cells have shown adipocyte inhibition of the ability of HSCs to proliferate and form hematopoietic colonies.[53]
Dysregulation
Cancer
Of the many transcriptional regulators of hematopoiesis, nearly all induce leukemia when aberrant. Chromosomal translocation is a hallmark of leukemia, and TAL1-induced translocation deregulates expression at the locus, while RUNX1-induced translocation results in chimeric fusion proteins. These chimeric transcription factors can result in the improper repression or activation of the target gene, as well as the inappropriate recruitment of chromatin-modifying enzymes.[65] PAX5 and Notch mutations can result in B-cell and T-cell leukemias, respectively.[66][67]. Dysregulation of stromal cells can in some cases induce genetic lesions in hematopoietic compartment; for example, mutations in the osteoblastic lineage cells resulted in malignant hematopoiesis [68][69]. Osteoblasts may also be dysregulated by the presence of solid tumors (outside the bone marrow); one study showed that mouse lung tumors increased osteoblast activity and numbers and that these cells were important to the outgrowth of the tumor in the lung via the production of tumor-infiltrating neutrophils [70].
Inflammation
Osteoblasts may be involved in other inflammatory systemic diseases, which is supported by studies using mouse models of sepsis[71]. Mesenchymal cells' response to β-adrenergic stimulation is altered in diabetes, which impairs G-CSF-induced HSCP mobilization.[72] Diabetes influences bone marrow endothelium, which may alter myeloid leukocyte generation[73][74]. This may be relevant for diabetes-associated morbidities such as atherosclerosis.[74]
References
- Birbrair, Alexander; Frenette, Paul S. (2016-03-01). "Niche heterogeneity in the bone marrow". Annals of the New York Academy of Sciences. 1370 (1): 82–96. Bibcode:2016NYASA1370...82B. doi:10.1111/nyas.13016. ISSN 1749-6632. PMC 4938003. PMID 27015419.
- Orkin SH (2000). "Diversification of haematopoietic stem cells to specific lineages". Nat. Rev. Genet. 1 (1): 57–64. doi:10.1038/35049577. PMID 11262875.
- Kim SI, Bresnick EH (2007). "Transcriptional control of erythropoiesis: emerging mechanisms and principles". Oncogene. 26 (47): 6777–6794. doi:10.1038/sj.onc.1210761. PMID 17934485.
- Galloway JL, Wingert RA, Thisse C, Thisse B, and Zon LI (2005). "Loss of gata1 but not gata2 converts erythropoiesis to myelopoiesis in zebrafish embryos". Dev. Cell. 8 (1): 109–116. doi:10.1016/j.devcel.2004.12.001. PMID 15621534.
- Rhodes J, Hagen A, Hsu K, Deng M, Liu TX, Look AT, and Kanki JP (2005). "Interplay of pu.1 and gata1 determines myelo-erythroid progenitor cell fate in zebrafish". Dev. Cell. 8 (1): 97–108. doi:10.1016/j.devcel.2004.11.014. PMID 15621533.
- Till J. E. & McCulloch E. (1961). "A direct measurement of the radiation sensitivity of normal mouse bone marrow cells". Radiat. Res. (Submitted manuscript). 14 (2): 213–222. Bibcode:1961RadR...14..213T. doi:10.2307/3570892. hdl:1807/2781. JSTOR 3570892. PMID 13776896.
- Schofield R. (1978). "The relationship between the spleen colony-forming cell and the haemopoietic stem cell". Blood Cells. 4 (1–2): 7–25. PMID 747780.
- Dexter T.M.; Allen T.D. & Lajha L.G. (1977). "Conditions controlling the proliferation of hemopoietic stem cells in vitro". J. Cell. Physiol. 91 (3): 335–344. doi:10.1002/jcp.1040910303. PMID 301143.
- Lord B.I.; Testa N.G.; Hendry J.H. (1975). "The relative spatial distributions of CFUs and CFUc in the normal mouse femur" (PDF). Blood. 46 (1): 65–72. doi:10.1182/blood.V46.1.65.65. PMID 1131427.
- Gong J.K. (1978). "Endosteal marrow: a rich source of hematopoietic stem cells". Science. 199 (4336): 1443–1445. Bibcode:1978Sci...199.1443G. doi:10.1126/science.75570. PMID 75570.
- Shalaby F, Ho J, Stanford WL, Fischer KD, Schuh AC, Schwartz L, Bernstein A, Rossant J (1997). "A requirement for Flk1 in primitive and definitive hematopoiesis and vasculogenesis". Cell. 89 (6): 981–990. doi:10.1016/S0092-8674(00)80283-4. PMID 9200616.
- Choi K, Kennedy M, Kazarov A, Papadimitriou JC, Keller G (1998). "A common precursor for hematopoietic and endothelial cells" (PDF). Development. 125 (4): 725–732. PMID 9435292.
- Ueno H, Weissman IL (2006). "Clonal analysis of mouse development reveals a polyclonal origin for yolk sac blood islands". Dev. Cell. 11 (4): 519–533. doi:10.1016/j.devcel.2006.08.001. PMID 17011491.
- Stainier DY, Weinstein BM, Detrich HW, third, Zon LI, Fishman MC (1995). "Cloche, an early acting zebrafish gene, is required by both the endothelial and hematopoietic lineages". Development. 121 (10): 3141–3150. PMID 7588049.
- Vogeli KM, Jin SW, Martin GR, Stainier DY (2006). "A common progenitor for haematopoietic and endothelial lineages in the zebrafish gastrula". Nature. 443 (7109): 337–339. Bibcode:2006Natur.443..337V. doi:10.1038/nature05045. PMID 16988712.
- Ema M, Rossant J (2003). "Cell fate decisions in early blood vessel formation". Trends Cardiovasc. Med. 13 (6): 254–259. doi:10.1016/S1050-1738(03)00105-1. PMID 12922023.
- Orkin SH, Zon LI (2008). "Hematopoiesis: An evolving paradigm for stem cell biology". Cell. 132 (4): 631–644. doi:10.1016/j.cell.2008.01.025. PMC 2628169. PMID 18295580.
- Wang LD, Wagers AJ (2011). "Dynamic niches in the origination and differentiation of haematopoietic stem cells". Nat. Rev. Mol. Cell Biol. 12 (10): 643–655. doi:10.1038/nrm3184. PMC 4040463. PMID 21886187.
- Kumano K, Chiba S, Kunisato A, Sata M, Saito T, et al. (2003). "Notch1 but not Notch2 is essential for generating hematopoietic stem cells from endothelial cells". Immunity. 18 (5): 699–711. doi:10.1016/S1074-7613(03)00117-1. PMID 12753746.
- Burns CE, Traver D, Mayhall E, Shepard JL, Zon LI (2005). "Hematopoietic stem cell fate is established by the Notch-Runx pathway". Genes Dev. 19 (19): 2331–2342. doi:10.1101/gad.1337005. PMC 1240042. PMID 16166372.
- Gering M, Patient R (2005). "Hedgehog signaling is required for adult blood stem cell formation in zebrafish embryos". Dev. Cell. 8 (3): 389–400. doi:10.1016/j.devcel.2005.01.010. PMID 15737934.
- Mascarenhas MI, Parker A, Dzierzak E, Ottersbach K (2009). "Identification of novel regulators of hematopoietic stem cell development through refinement of stem cell localization and expression profiling". Blood. 114 (21): 4645–4653. doi:10.1182/blood-2009-06-230037. PMC 2780301. PMID 19794138.
- Esner M; et al. (2006). "Smooth muscle of the dorsal aorta shares a common clonal origin with skeletal muscle of the myotome". Development. 133 (4): 737–749. doi:10.1242/dev.02226. PMID 16436625.
- Ma Q; et al. (1998). "Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice". Proc Natl Acad Sci USA. 95 (16): 9448–9453. Bibcode:1998PNAS...95.9448M. doi:10.1073/pnas.95.16.9448. PMC 21358. PMID 9689100.
- Mcgrath KE, Koniski AD, Maltby KM, et al. (1999). "Embryonic expression and function of the chemokine SDF-1 and its receptor, CXCR4". Develop Biol. 213 (2): 442–456. doi:10.1006/dbio.1999.9405. PMID 10479460.
- Christensen JL, Wright DE, Wagers AJ, Weissman IL (2004). "Circulation and chemotaxis of fetal hematopoietic stem cells". PLoS Biol. 2 (3): e75. doi:10.1371/journal.pbio.0020075. PMC 368169. PMID 15024423.
- Broxmeyer HE; et al. (1991). "The kit receptor and its ligand, steel factor, as regulators of hematopoiesis". Cancer Cells. 3 (12): 480–487. PMID 1726456.
- Qian H; et al. (2007). "Distinct roles of integrins α6 and α4 in homing of fetal liver hematopoietic stem and progenitor cells". Blood. 110 (7): 2399–2407. doi:10.1182/blood-2006-10-051276. PMID 17586725.
- Martin, MA, Bhatia, M (2005). "Analysis of the human fetal liver hematopoietic microenvironment". Stem Cells and Development. 14 (5): 493–504. doi:10.1089/scd.2005.14.493. PMID 16305335.
- Kieusseian A; et al. (2006). "Expression of Pitx2 in stromal cells is required for normal hematopoiesis". Blood. 107 (2): 492–500. doi:10.1182/blood-2005-02-0529. PMC 1895608. PMID 16195330.
- Shiojiri, N. (1997). "Development and differentiation of bile ducts in the mammalian liver". Microsc. Res. Tech. 39 (4): 328–335. doi:10.1002/(SICI)1097-0029(19971115)39:4<328::AID-JEMT3>3.0.CO;2-D. PMID 9407543.
- Inman KE, Downs KM (2007). "The murine allantois: emerging paradigms in development of the mammalian umbilical cord and its relation to the fetus". Genesis. 45 (5): 237–258. doi:10.1002/dvg.20281. PMID 17440924.
- Wolber, FM; et al. (2002). "Roles of spleen and liver in development of the murine hematopoietic system". Exp. Hematol. 30 (9): 1010–1019. doi:10.1016/S0301-472X(02)00881-0. PMID 12225792.
- Chan C; et al. (2009). "Endochondral ossification is required for haematopoietic stem-cell niche formation". Nature. 457 (7228): 490–494. Bibcode:2009Natur.457..490C. doi:10.1038/nature07547. PMC 264814. PMID 19078959.
- Magnon C, Frenette PS (2008). "Hematopoietic stem cell trafficking". Stembook. doi:10.3824/stembook.1.8.1. PMID 20614595. Missing or empty
|title=(help) - Broxmeyer HE; et al. (2005). "Rapid mobilization of murine and human hematopoietic stem and progenitor cells with AMD3100, a CXCR4 antagonist". J. Exp. Med. 201 (8): 1307–1318. doi:10.1084/jem.20041385. PMC 2213145. PMID 15837815.
- Kim I, Saunders TL, Morrison SJ (2007). "Sox17 dependence distinguishes the transcriptional regulation of fetal from adult hematopoietic stem cells". Cell. 130 (3): 470–483. doi:10.1016/j.cell.2007.06.011. PMC 2577201. PMID 17655922.
- Ichikawa M, Asai T, Saito T, et al. (2004). "AML-1 is required for megakaryocytic maturation and lymphocytic differentiation, but not for maintenance of hematopoietic stem cells in adult hematopoiesis". Nat. Med. 10 (3): 299–304. doi:10.1038/nm997. PMID 14966519.
- Parmar K, Mauch P, Vergilio JA, et al. (2007). "Distribution of hematopoietic stem cells in the bone marrow according to regional hypoxia". Proc. Natl. Acad. Sci. USA. 104 (13): 5431–5436. Bibcode:2007PNAS..104.5431P. doi:10.1073/pnas.0701152104. PMC 1838452. PMID 17374716.
- Winkler IG; et al. (2010). "Positioning of bone marrow hematopoietic and stromal cells relative to blood flow in vivo: serially reconstituting hematopoietic stem cells reside in distinct nonperfused niches". Blood. 116 (3): 375–385. doi:10.1182/blood-2009-07-233437. PMID 20393133.
- Boitano AE; et al. (2010). "Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells". Science. 329 (5997): 1345–1348. Bibcode:2010Sci...329.1345B. doi:10.1126/science.1191536. PMC 3033342. PMID 20688981.
- Simsek T; et al. (2010). "The distinct metabolic profile of hematopoietic stem cells reflects their location in a hypoxic niche". Cell Stem Cell. 7 (3): 380–390. doi:10.1016/j.stem.2010.07.011. PMC 4159713. PMID 20804973.
- Takubo K; et al. (2010). "Regulation of the HIF-1α level is essential for hematopoietic stem cells". Cell Stem Cell. 7 (3): 391–402. doi:10.1016/j.stem.2010.06.020. PMID 20804974.
- Adams GB; et al. (2006). "Stem cell engraftment at the endosteal niche is specified by the calcium-sensing receptor". Nature. 439 (7076): 599–603. Bibcode:2006Natur.439..599A. doi:10.1038/nature04247. PMID 16382241.
- Lam BS, Cunningham C, Adams GB (2011). "Pharmacologic modulation of the calcium-sensing receptor enhances hematopoietic stem cell lodgment in the adult bone marrow". Blood. 117 (4): 1167–1175. doi:10.1182/blood-2010-05-286294. PMC 3056470. PMID 21076044.
- Lymperi S, Ersek A, Ferraro F, Dazzi F, Horwood NJ (2011). "Inhibition of osteoclast function reduces hematopoietic stem cell numbers in vivo". Blood. 117 (5): 1540–1549. doi:10.1182/blood-2010-05-282855. PMID 21131587.
- Adamo L; et al. (2009). "Biomechanical forces promote embryonic haematopoiesis". Nature. 459 (7250): 1131–1135. Bibcode:2009Natur.459.1131A. doi:10.1038/nature08073. PMC 2782763. PMID 19440194.
- Keung AJ, Healy KE, Kumar S, Schaffer DV (2010). "Biophysics and dynamics of natural and engineered stem cell microenvironments" (PDF). Wiley Interdiscip. Rev. Syst. Biol. Med. 2 (1): 49–64. doi:10.1002/wsbm.46. PMID 20836010.
- Engler AJ, Sen S, Sweeney HL, Discher DE (2006). "Matrix elasticity directs stem cell lineage specification". Cell. 126 (4): 677–689. doi:10.1016/j.cell.2006.06.044. PMID 16923388.
- Gilbert PM; et al. (2010). "Substrate elasticity regulates skeletal muscle stem cell self-renewal in culture". Science. 329 (5995): 1078–1081. Bibcode:2010Sci...329.1078G. doi:10.1126/science.1191035. PMC 2929271. PMID 20647425.
- Calvi LM; et al. (2003). "Osteoblastic cells regulate the haematopoietic stem cell niche". Nature. 425 (6960): 841–846. Bibcode:2003Natur.425..841C. doi:10.1038/nature02040. PMID 14574413.
- Visnjic, D (2004). "Hematopoiesis is severely altered in mice with an induced osteoblast deficiency". Blood. 103 (9): 3258–3264. doi:10.1182/blood-2003-11-4011. PMID 14726388.
- Chitteti BR; et al. (2010). "Impact of interactions of cellular components of the bone marrow microenvironment on hematopoietic stem and progenitor cell function". Blood. 115 (16): 3239–3248. doi:10.1182/blood-2009-09-246173. PMC 2858485. PMID 20154218.
- Nakamura Y; et al. (2010). "Isolation and characterization of endosteal niche cell populations that regulate hematopoietic stem cells". Blood. 116 (9): 1422–1432. doi:10.1182/blood-2009-08-239194. PMID 20472830.
- Kohler A; et al. (2009). "Altered cellular dynamics and endosteal location of aged early hematopoietic progenitor cells revealed by time-lapse intravital imaging in long bones". Blood. 114 (2): 290–298. doi:10.1182/blood-2008-12-195644. PMC 2714205. PMID 19357397.
- Lo Celso C; et al. (2009). "Live-animal tracking of individual haematopoietic stem/progenitor cells in their niche". Nature. 457 (7225): 92–97. Bibcode:2009Natur.457...92L. doi:10.1038/nature07434. PMC 2820276. PMID 19052546.
- Xie Y; et al. (2009). "Detection of functional haematopoietic stem cell niche using real-time imaging". Nature. 457 (7225): 97–101. Bibcode:2009Natur.457...97X. doi:10.1038/nature07639. PMID 19052548.
- Méndez-Ferrer S; et al. (2010). "Mesenchymal and haematopoietic stem cells form a unique bone marrow niche". Nature. 466 (7308): 829–834. Bibcode:2010Natur.466..829M. doi:10.1038/nature09262. PMC 3146551. PMID 20703299.
- Tzeng YS; et al. (2011). "Loss of Cxcl12/Sdf-1 in adult mice decreases the quiescent state of hematopoietic stem/progenitor cells and alters the pattern of hematopoietic regeneration after myelosuppression". Blood. 117 (2): 429–439. doi:10.1182/blood-2010-01-266833. PMID 20833981.
- Katayama Y; et al. (2006). "Signals from the sympathetic nervous system regulate hematopoietic stem cell egress from bone marrow". Cell. 124 (2): 407–421. doi:10.1016/j.cell.2005.10.041. PMID 16439213.
- Omatsu Y; et al. (2010). "The essential functions of adipoosteogenic progenitors as the hematopoietic stem and progenitor cell niche". Immunity. 33 (3): 1–13. doi:10.1016/j.immuni.2010.08.017. PMID 20850355.
- Hooper AT; et al. (2009). "Engraftment and reconstitution of hematopoiesis is dependent on VEGFR2-mediated regeneration of sinusoidal endothelial cells". Cell Stem Cell. 4 (3): 263–274. doi:10.1016/j.stem.2009.01.006. PMC 3228275. PMID 19265665.
- Kobayashi H; et al. (2010). "Angiocrine factors from Akt-activated endothelial cells balance self-renewal and differentiation of haematopoietic stem cells". Nature Cell Biology. 12 (11): 1046–1056. doi:10.1038/ncb2108. PMC 2972406. PMID 20972423.
- Naveiras O; et al. (2009). "Bone-marrow adipocytes as negative regulators of the haematopoietic microenvironment". Nature. 460 (7252): 259–263. Bibcode:2009Natur.460..259N. doi:10.1038/nature08099. PMC 2831539. PMID 19516257.
- Rosenbauer F, Tenen DG (2007). "Transcription factors in myeloid development: balancing differentiation with transformation". Nat. Rev. Immunol. 7 (2): 105–117. doi:10.1038/nri2024. PMID 17259967.
- Mullighan CG, Goorha S, Radtke I, et al. (2007). "Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia". Nature. 446 (7137): 758–764. Bibcode:2007Natur.446..758M. doi:10.1038/nature05690. PMID 17344859.
- Weng AP, Ferrando AA, Lee W, et al. (2004). "Activating mutations of NOTCH1 in human T cell acute lymphoblastic leukemia". Science. 306 (5694): 269–271. Bibcode:2004Sci...306..269W. CiteSeerX 10.1.1.459.5126. doi:10.1126/science.1102160. PMID 15472075.
- Raaijmakers, Marc H. G. P.; Mukherjee, Siddhartha; Guo, Shangqin; Zhang, Siyi; Kobayashi, Tatsuya; Schoonmaker, Jesse A.; Ebert, Benjamin L.; Al-Shahrour, Fatima; Hasserjian, Robert P.; Scadden, Edward O.; Aung, Zinmar; Matza, Marc; Merkenschlager, Matthias; Lin, Charles; Rommens, Johanna M.; Scadden, David. T. (21 March 2010). "Bone progenitor dysfunction induces myelodysplasia and secondary leukaemia". Nature. 464 (7290): 852–857. Bibcode:2010Natur.464..852R. doi:10.1038/nature08851. PMC 3422863. PMID 20305640.
- Kode, Aruna; Manavalan, John S.; Mosialou, Ioanna; Bhagat, Govind; Rathinam, Chozha V.; Luo, Na; Khiabanian, Hossein; Lee, Albert; Murty, Vundavalli V.; Friedman, Richard; Brum, Andrea; Park, David; Galili, Naomi; Mukherjee, Siddhartha; Teruya-Feldstein, Julie; Raza, Azra; Rabadan, Raul; Berman, Ellin; Kousteni, Stavroula (15 January 2014). "Leukaemogenesis induced by an activating β-catenin mutation in osteoblasts". Nature. 506 (7487): 240–244. Bibcode:2014Natur.506..240K. doi:10.1038/nature12883. PMC 4116754. PMID 24429522.
- Engblom, Camilla; Pfirschke, Christina; Zilionis, Rapolas; Da Silva Martins, Janaina; Bos, Stijn A.; Courties, Gabriel; Rickelt, Steffen; Severe, Nicolas; Baryawno, Ninib; Faget, Julien; Savova, Virginia; Zemmour, David; Kline, Jaclyn; Siwicki, Marie; Garris, Christopher; Pucci, Ferdinando; Liao, Hsin-Wei; Lin, Yi-Jang; Newton, Andita; Yaghi, Omar K.; Iwamoto, Yoshiko; Tricot, Benoit; Wojtkiewicz, Gregory R.; Nahrendorf, Matthias; Cortez-Retamozo, Virna; Meylan, Etienne; Hynes, Richard O.; Demay, Marie; Klein, Allon; Bredella, Miriam A.; Scadden, David T.; Weissleder, Ralph; Pittet, Mikael J. (1 December 2017). "Osteoblasts remotely supply lung tumors with cancer-promoting SiglecF high neutrophils". Science. 358 (6367): eaal5081. doi:10.1126/science.aal5081. PMC 6343476. PMID 29191879.
- Terashima, Asuka; Okamoto, Kazuo; Nakashima, Tomoki; Akira, Shizuo; Ikuta, Koichi; Takayanagi, Hiroshi (June 2016). "Sepsis-Induced Osteoblast Ablation Causes Immunodeficiency". Immunity. 44 (6): 1434–1443. doi:10.1016/j.immuni.2016.05.012. PMID 27317262.
- Ferraro, F.; Lymperi, S.; Mendez-Ferrer, S.; Saez, B.; Spencer, J. A.; Yeap, B. Y.; Masselli, E.; Graiani, G.; Prezioso, L.; Rizzini, E. L.; Mangoni, M.; Rizzoli, V.; Sykes, S. M.; Lin, C. P.; Frenette, P. S.; Quaini, F.; Scadden, D. T. (12 October 2011). "Diabetes Impairs Hematopoietic Stem Cell Mobilization by Altering Niche Function". Science Translational Medicine. 3 (104): 104ra101–104ra101. doi:10.1126/scitranslmed.3002191. PMC 3754876.
- Mangialardi, Giuseppe; Katare, Rajesh; Oikawa, Atsuhiko; Meloni, Marco; Reni, Carlotta; Emanueli, Costanza; Madeddu, Paolo (March 2013). "Diabetes Causes Bone Marrow Endothelial Barrier Dysfunction by Activation of the RhoA–Rho-Associated Kinase Signaling Pathway". Arteriosclerosis, Thrombosis, and Vascular Biology. 33 (3): 555–564. doi:10.1161/ATVBAHA.112.300424.
- Hoyer, FF; Zhang, X; Coppin, E; Vasamsetti, SB; Modugu, G; Schloss, MJ; Rohde, D; McAlpine, CS; Iwamoto, Y; Libby, P; Naxerova, K; Swirski, FK; Dutta, P; Nahrendorf, M (22 April 2020). "Bone Marrow Endothelial Cells Regulate Myelopoiesis in Diabetes". Circulation. doi:10.1161/CIRCULATIONAHA.120.046038. PMID 32316750.
| Sources/types | |
|---|---|
| Cell potency |
|
| Related articles |
|
| |