Drosophila subobscura
Drosophila subobscura is a species of fruit fly in the family Drosophilidae. Originally found around the Mediterranean, it has spread to most of Europe and the Near East. It has been introduced into the west coasts of Canada, the United States, and Chile. Its closest relative is Drosophila madeirensis, found in the Madeira Islands, followed by D. guanche, found in the Canary Islands. These three species form the D. subobscura species subgroup. When they mate, males and females perform an elaborate courtship dance, in which the female can either turn away to end the mating ritual, or stick out her proboscis in response to the male's, allowing copulation to proceed. D. subobscura has been regarded as a model organism for its use in evolutionary-biological studies.
| Drosophila subobscura | |
|---|---|
_subobscura_(male)_collage.jpg) | |
| Five anatomical images of a Drosophila subobscura (male) on a rotting squash, located near Christchurch Park, Ipswich | |
| Scientific classification | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | |
| Family: | |
| Genus: | |
| Subgenus: | |
| Species group: | obscura |
| Species subgroup: | subobscura |
| Species: | D. subobscura |
| Binomial name | |
| Drosophila subobscura Collin, 1936 | |
Description
Both wild type and laboratory-reared individuals of D. subobscura are brown, with clear wings, yellow halters, yellowish legs, and red eyes. They do not exhibit sexual size dimorphism; the males and females are about 2 mm (0.08 in) long. The males have a brown antenna, grey pollinose, and brown dorsal surface of the thorax. The tergum (dorsal area excluding the head) is dark brown. There are three tarsal segments on the fore legs. The first segment is the longest and contains the proximal comb, which has 7-12 teeth. The second segment contains the distal comb, which has 10-13 teeth. The females share the same characteristics as the males, except in respect to tarsal combs. The eggs are only half the length of those of the closely related D. obscura species. Its larvae and pupa are of the usual drosophilid type.[1]
Head
The arista (bristles arising from the antennae) of D. subobscura contain 6-8 branches, with 1-2 of those branches below the terminal fork. The species has a brown antenna with grey pollinosity that is approximately the same color as the rest of the head-capsule. The front of the antenna is dark brown and matte, without any pollinosity except on the frontal triangle and fronto-orbital plates, both of which are shiny and slightly pollinose. The carina (tracheal cartilage that divides the two bronchi) of the fly is rounded, widening below, and the face is a paler brown color with grey pollinosoty. The genae is brown with heavy, grey pollinosity. The ocelli (small or 'simple' eyes of an insect) are the same color as the eyes.[1]

Body
The dorsa of the thorax is brown with grey pollinose. There appear to be no traces of any longitudinal stripes or lines upon it.[1] The wings are colorless. Their membrane never folds nor crumples, but instead expands and displays intricate venation.[1][2]
_(18166344402).jpg)
The legs of D. subobscura are a yellowish color. The combs of the teeth are each aligned on the longitudinal axis of the first and second tarsal segments of the fore legs. The proximal comb, identified as the comb of the first segment, has 7-12 teeth and is about half as long as the segment itself. Teeth on the distal end of the segment are slightly longer than the segment. The first tarsal segment is slightly longer than both the second and third segments, but shorter than their combined length. The abdomen has tergites that are uniformly dark brown, but shiny in some lights. Other lights reveal a grey pollinosity appearance.[1]
Bristles
The upper reclinate fronto-orbital bristles are long, the middle reclinate bristles are short, and the lower proclinate bristles are medium in length. Proportionally, the listed bristles lengths can be characterized in a 4:2:3 fashion. The ocellar, post-vertical, and inner and outer vertical bristles are all about the same length as the upper reclinate fronto-orbital bristles. The bristle behind the vibrissa is less than half the length of the upper reclinate fronto-orbital bristles. The species also has two pairs of dorso-central bristles, which contain 8-10 rows of acrostichal hairs. No acrostichal bristles are seen to have developed in the species. The anterior scutella bristles are parallel to each other. The anterior sternopleural bristles are shorter than the posterior ones. The wings have costal bristles. The pre-apical tibial and apical tibial bristles of the legs are not exceptionally long.[1]
Distinction from other species of Drosophila
D. subobscura are found to be unusual among the Drosophila genus, because they are monandrous (females only mate one at a time).[3][4] Additionally, unlike the rest of the Drosophila genus, D. subobscura do not mate in the absence of light[5][6] nor do they produce courtship songs by wing vibration.[7] A study published in 2017 revealed that the difference in courtship behavior between D. subobscura and D. melanogaster (in particular, the D. subobscura's nuptial gift transfer behavior) could be potentially due to the optogenetic activation of a distinct neural circuit that differs in both flies.[8]
Taxonomy
In 1933, A. H. Sturtevant captured a species of Drosophila in England. When Sturtevant submitted the captured specimens to J. E. Collin of Newmarket, Collin initially misidentified the species as D. obscura. Three years later, the first description of D. subobscura appeared in an addendum to Gordon's paper through a short note written by Collin. In the note, Collin compared both sexes of D. subobscura and differentiated them as a separate species from D. obscura, their nearest related species. Collin's description, considered incomplete but necessary to validate the D. subobscura name, was followed up with a more complete description by Dr. James Smart.[9][10][1]
In 1942, A. H. Sturtevant founded the obscura group, which initially consisted of the affinis and obscura species groups.[11] The obscura group falls under the subgenus Sophophora. The obscura species group currently contains 6 subgroups, listed alphabetically: affinis, microlabis, obscura, pseudoobscura, subobscura, and sinobscura. D. subobscura belongs to the subobscura subgroup, along with the closely related Drosophila guanche and Drosophila madeirensis.[12]
In experimental trials, D. subobscura does not breed with any other species of obscura, except D. madeirensis, a species also in the subobscura subgroup. When crossed, sterile males and fertile female hybrids are formed.[13] As of March 2019, the first long-read sequencing of D. subobscura’s genome has been presented, showing that evolution of its genome structure is indirectly driven by the effects of suppressing recombination of genetic inversions. This suppressive effect maintains various sets of adaptive alleles together in the midst of gene flow.[14]
When Collin identified Sturtevant's captured species as D. subobscura in 1933, the name was coined as a manuscript name. The species was then bred at the Department of Biometry in University College in London to be genetically experimented on. Here, one paper that resulted from this experimental work in London referred to the tested species as Collin's coined manuscript name, D. subobscura, and was referred to as such moving forward. During this time, involved parties knew that D. subobscura was simply a manuscript name, thus creating anticipation that Collin would publish a description of the species and consequently validate the name. Although Collin did not publish the anticipated description of the species, in 1936, he contributed a note in the addendum of a paper published by Gordon that same year. The note outlines a diagnosis of the sexes and differentiation of the species from D. obscura and attributes the D. subobscura name to Collin. Thus, the name “D. subobscura Collin” dates from 1936, because none of the papers that come before it include a description of the given species that would have satisfied nomenclature rules. Therefore, prior to the addendum in Gordon's paper in 1936, the D. subobscura name used in published works must be regarded as a manuscript name. In 1938, two years after Gordon's paper, Dr. Eugéne Séguy had discovered a new species of Drosophila in Kenya, naming it D. subobscura. Though this D. subobscura was recorded in the 1938 Zoological Record, the “D. subobscura Collin” name has not appeared in the Zoological Record as a new species, yet.[1][15][16][17]
Distribution
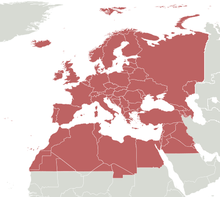
D. subobscura is widely distributed in Europe, from Scandinavia south to the Mediterranean, and in North Africa and the Middle East as far east as Iran.[18] Its distribution spans over thirty latitudinal degrees, with its most dense populations residing in the western Palaeartic realm.[19] Introduced populations of D. subobscura are found in the west coasts of Canada, the United States, and Chile.[20]
D. subobscura was first discovered in the Americas (southern Chile) in February 1978.[21] D. subobscura was later found in La Serena, Chile, in the summer of 1979;[22] Punta Arenas, Chile, in January 1981; San Carlos de Bariloche, Argentina, in November 1981; and then Mar de Plata, Argentina, in 1984. In 1982, D. subobscura was discovered in North America in the city of Port Townsend, Washington, followed by the surrounding northern and southern areas, from Vancouver B.C. to Oregon. In fall 1983, D. subobscura was found in the Central Valley, Davis, and El Rio areas of California.[23]
There has been recent speculation about D. subobscura colonization in western North America being a more modern-event. The origin of the North and South America colonizers remains unknown, but evidence reveals that they derive from the original, Palearctic populations.[20]
Habitat

D. subobscura is mainly found in open fields or forest fringes.[24] Decreases in light and temperature induce locomotion activity in D. subobscura towards areas outside of the forest.[25] The genome of some Greek populations of D. subobscura has shown evidence of microgeographic variation, prompting a possibility that the species exhibits habitat choice.[26]
However, no evidence has been found to show that D. subobscura exhibits individual habitat choice, aligning with the fact that its well-studied inversion polymorphism is relatively inflexible and slow to respond to the environment. Additionally, individuals do not exhibit much preference for different times of the day; even then, there remains some evidence to suggest that the species displays feeding and breeding site fidelity, as individuals were shown to return to familiar baits.[27]
Mating

D. subobscura is monandrous, a behavior not usually seen among Drosophila.[28] Visual stimuli dictate courtship behavior.[29] D. subobscura do not mate in the dark[30] and do not produce a courtship song via wing vibrations like other species of Dipterans.[31] Instead, the lone male repeatedly “scissors” its wings, an activity augmented in the presence of other flies. This behavior indicates that the male is looking to court a female. When a female appears, the male taps her with his own front legs. The male then stands in front and directly faces the female to stick out his proboscis. The male and female then start to “dance”, as the female rapidly sidesteps, while the male tries to keep himself directly facing the female. During the dance, the male's wings are usually raised and extended. Sometimes, mounting can occur without a dance taking place prior. At this point, the female has two options: she can either end the dance, without mating, by turning away from the male and leaving, or she can stand still, extend her own proboscis, and invite the male to mount her by parting her wings. In the latter behavior, the male stretches its wings sideways and swings behind the female to mount her. The male and female probosces may or may not touch beforehand.[29] Additionally, unique to just D. subobscura among the other species in its subgroup, males will attempt to mate with wax models, only if the wax is moved in the patterns similar to female-male courtship dances. If wax models did not carry out the dance, then ultimately the males did not attempt copulation.[32]
Although males always extend their own proboscis, this activity within the female varies greatly. Activities among three consecutive male-female courtships showed three different female behaviors: 1) no protrusion of the proboscis, 2) continued extension of the proboscis for several seconds after mounting had occurred, and 3) repeated protrusion and withdrawal of the proboscis before the male mounted.[29] The tips of the male and female probosces can be observed to be brought into contact, where they alternate with back and forth motions.[33]
Sexual selection
D. subobscura practice nuptial feeding, a practice where a nutritional gift is transferred from one partner to another during/directly after courtship and/or copulation. In the case of D. subobscura, the gift is a regurgitated drop of liquid secreted from the male's crop, onto the female's proboscis.[34] Preventing production and exchange of nutritional gifts among D. subobscura has been shown to decrease both male mating success and egg count among females.[35]
It has been shown that males that are in good condition produce more nutritional gifts, thereby increasing their mating success. Additionally, starved females show preference for well-fed males as a way to increase the female's fecundity – this preference is speculated to be the case due to the greater quantity of drops that well-fed males produce.[36] If larger males, carrying bigger nutritional gifts, are prevented from producing their gifts, then small males are more successful in female courtship, due to better tracking of the female during the courtship dance. Larger males are seen to have slower acceleration and deceleration speeds.[37]
Inbred male behavior
Reported observations of mating behavior in inbred males reveal that in most cases, active courtships were seen, but mating did not normally follow. However, prolonged dances were rare. In some instances, the male directly approaches the female, in which the female steps sideways several times in front of the stationary male, before ultimately turning away. Occasionally, the male attempts to follow the sidestep movements of the female but would often lag behind and struggle to consistently face the female. In the incident that the female stands still and extends her proboscis, the male usually would attempt to mount. More often than not, the inbred male would fall on his back, or land too far forward or too far back on the female. In the case of the latter, the female normally stands still with her wings partially extended before eventually kicking off the male. Inbred males who have continuously but unsuccessfully attempted to court a female may approach the female from the side or behind and attempt to directly mount, a behavior described as “desperation” to some scientists. These mating attempts remain unsuccessful. The observed lower mating success in inbred males has been thought to be due to lower athletic ability via physiologically-efficient muscles, sense organs, and neuromuscular coordination, rather than lower intensity of courtship.[29]
Influence of gut microbiota on mating behavior
A study has displayed that the condition of D. subobscura's gut microbiota can have an effect on its mating behavior.[38] Upon suppressing the gut bacteria of female D. subobscura with antibiotics, researchers observed that these females mated faster with males that had intact microbiota. Females with intact gut bacteria were less willing to mate with males that had intact microbiota. Additionally, fecundity was seen to increase when the gut bacteria of male and female D. subobscura were suppressed through antibiotics, compared to no suppression.[38]
Genetics
Analysis of D. subobscura’s salivary gland has shown that its genome mimics the Drosophila karyotype, consisting of a small dot and five large acrocentric rods. Additionally, the genome does not show a chromocenter and contains high levels of chromosomal polymorphisms caused by paracentric inversions on all of the acrocentric rods.[39] Polytene drawings and photomaps helped further the study of these inversions, allowing for the finding of more than 600 different linkages and genetic markers, which encompass a majority of the euchromatic genome.[40][41][42][43] More than 65 inversions have been identified.[44]
D. subobscura is frequently used in evolutionary-biological studies.[45][46][47][48][49] As D. subobscura, among others within its species group, has been reputed as a model organism for evolutionary-biological studies, its genetics and ecology have been scrutinized for more than forty years.[50] These flies have served as favorable models ever since Theodosius Dobzhansky and his colleagues published their influential works in the 1930s.[51][52] From the species' discovery in the Palearctic realm to its colonization of North and South America, it has attracted the interests of both European and American scientists as experimental material in evolution, biology, and ecology.[53]
Significance in tracking climate change
The D. subobscura genome has been used to track global climate change by measuring the magnitude and direction of shifts in chromosome inversion frequencies in comparison to ambient temperatures at selected European, North American. and South American sites.[54] In 21 of 22 populations of D. subobscura, genotypes seen in warm climates increased in frequency. It was shown that genetic changes in D. subobscura at these sites can be used as a possible tool to track global climate warming.[54]
References
- "Smart, J. 1945. Drosophila subobscura Collin: descriptive notes on the species with comments on its nomenclatorial status (Diptera). Proc. R. Entomol. Soc. Lond. Ser. B. Taxon, 14:53-56" (PDF). Retrieved 24 September 2019.
- Gordon, C.; Spurway, H.; Street, P. A. R. (1939). "An analysis of three wild populations of Drosophila subobscura". J. Genet. 38 (1–2): 37–90. doi:10.1007/BF02982165.
- Smith, J. Maynard (1956-05-01). "Fertility, mating behaviour and sexual selection in Drosophila Subobscura". Journal of Genetics. 54 (2): 261–279. doi:10.1007/BF02982781. ISSN 0022-1333.
- Fisher, David N.; Doff, Rowan J.; Price, Tom A. R. (2013-07-25). "True polyandry and pseudopolyandry: why does a monandrous fly remate?". BMC Evolutionary Biology. 13 (1): 157. doi:10.1186/1471-2148-13-157. ISSN 1471-2148. PMC 3728105. PMID 23885723.
- Philip, U.; Rendel, J. M.; Spurway, H.; Haldane, J. B. S. (1944-08-01). "Genetics and Karyology of Drosophila Subobscura". Nature. 154 (3904): 260–262. Bibcode:1944Natur.154..260P. doi:10.1038/154260a0. ISSN 1476-4687.
- Rendel, J. M. (1945-01-01). "Genetics and cytology of Drosophila subobscura". Journal of Genetics. 46 (2): 287–302. doi:10.1007/BF02989271. ISSN 0022-1333.
- Ewing, Arthur W.; Bennet-Clark, H. C. (1968-01-01). "The Courtship Songs of Drosophila". Behaviour. 31 (3–4): 288–301. doi:10.1163/156853968X00298. ISSN 0005-7959.
- Tanaka, Ryoya; Higuchi, Tomohiro; Kohatsu, Soh; Sato, Kosei; Yamamoto, Daisuke (2017-11-29). "Optogenetic Activation of the fruitless-Labeled Circuitry in Drosophila subobscura Males Induces Mating Motor Acts". The Journal of Neuroscience. 37 (48): 11662–11674. doi:10.1523/JNEUROSCI.1943-17.2017. ISSN 0270-6474. PMC 6705751. PMID 29109241.
- Gordon, C (1936). "The frequency of heterozygous in free-living populations of Drosophila melanogaster and Drosophila subobscura". J. Genet.: 25–33.
- Collin, J. E. (1936). "Note Drosophila subobscura sp.n. male, female. Addendum to Gordon". J. Genet.: 60.
- "A. H. Sturtevant 1942. The classification of the genus Drosophila with the description of nine new species. Univ Texas Publ 4213:5–51" (PDF). Retrieved 24 September 2019.
- "Therese A. Markow; Patrick M. O'Grady (2005). Drosophila: A guide to species identification and use. London: Elsevier. ISBN 978-0-12-473052-6". Retrieved 24 September 2019.
- Khadem, M.; Krimbas, C. B. (1991). "Studies of the species barrier between Drosophila subobscura and D. madeirensis. IV. A genetic dissection of the X chromosome for speciation gene". Hereditas. 114 (2): 189–95. doi:10.1111/j.1601-5223.1991.tb00323.x. PMID 1917545.
- Karageorgiou, Charikleia; Gámez-Visairas, Víctor; Tarrío, Rosa; Rodríguez-Trelles, Francisco (2019). "Long-read based assembly and synteny analysis of a reference Drosophila subobscura genome reveals signatures of structural evolution driven by inversions recombination-suppression effects". BMC Genomics. 20 (1): 223. doi:10.1186/s12864-019-5590-8. PMC 6423853. PMID 30885123.
- Drosophila Information Service (1939–43). Nos. 11–17. Reports from Biometry Department, University College, London.
- Séguy, Drosophila subobscura, 1938, Mem. Mjs. Hist. nat. Paris (n.s.) 8:352.
- Séguy, Drosophila subobscura, 1939, Zool. Rec. 75 (Pt. 12):384.
- "Bächli, G. "Quantitative methods for population analysis applied to a Drosophila (Diptera, Drosophilidae) collection." Aquilo (Zool) 20 (1979): 33-40". Cite journal requires
|journal=(help) - "Buzzati-Traverso AA, Scossiroli RE. The "obscura group" of the genus Drosophila. Adv Genet. 1955;7:47–92". Retrieved 24 September 2019.
- Ayala, Francisco J.; Serra, Luis; Prevosti, Antonio (1989). "A grand experiment in evolution: The Drosophila subobscura colonization of the Americas". Genome. 31: 246–255. doi:10.1139/g89-042. S2CID 55247939.
- Budnik, M; Benado, M; Cifuentes, L. (1996). "Colonization of Drosophila subobscura in Chile: Experimental preadult competition with local species under bi- and tri-specific conditions". Netherlands Journal of Zoology. 47 (2): 133–141. doi:10.1163/156854297X00148. S2CID 84917494.CS1 maint: multiple names: authors list (link)
- Prevosti A.; Serra L.; et al. (1989). "Colonization and establishment of the Palearctic species Drosophila subobscura in North and South America". Evolutionary Biology of Transient Unstable Populations (ed. Fontdevila A.). Springer-Verlag, Berlin. pp. 114–129. doi:10.1007/978-3-642-74525-6_8. ISBN 978-3-642-74527-0.
- Beckenbach, Andrew T.; Prevosti, Antonio (1986). "Colonization of North America by the European Species, Drosophila subobscura and D. ambigua". American Midland Naturalist. 115 (1): 10–18. doi:10.2307/2425832. JSTOR 2425832.
- "Burla, H.: Systematik, Verbreitung und Ökologie der Drosophila-Arten der Schweiz. Rev. suisse Zool. 58, 23–175 (1951)". Cite journal requires
|journal=(help) - Koch, Rudolf (1967). "Tagesperiodik der Aktivität und der Orientierung nach Wald und Feld von Drosophila subobscura und Drosophila obscura". Zeitschrift für Vergleichende Physiologie. 54 (3): 353–394. doi:10.1007/BF00298224.
- Krimbas, C.B.; Alevizos, V. (1973). "The genetics of Drosophila subobscura populations. IV. Further data on inversion polymorphism in Greece—Evidence of microdifferentiation". Egypt. J. Genet. Cytol. 2 (12): 1–132.
- Atkinson, W. D.; Miller, J. A. (1980). "Lack of habitat choice in a natural population of Drosophila subobscura". Heredity. 44 (2): 193–199. doi:10.1038/hdy.1980.16.
- Holman, L.; Freckleton, R. P.; Snook, R. R. (2008). "What use is an infertile sperm? A comparative study of sperm-heteromorphic Drosophila". Evolution. 62 (2): 374–85. doi:10.1111/j.1558-5646.2007.00280.x. PMID 18053077.
- Maynard Smith, J. (1956). "Fertility, mating behaviour and sexual selection in Drosophila subobscura". Journal of Genetics. 54 (2): 261–279. doi:10.1007/BF02982781. PMID 15876580.
- Rendel, J. M. (1945). "Genetics and cytology of D. subobscura. II. Normal and selective matings in Drosophila subobscura". J. Genet. 46: 287–302. doi:10.1007/BF02989271.
- Ewing, Arthur W.; Bennet-Clark, H.C. (1968). "The courtship songs of Drosophila". Behaviour. 31 (3–4): 288–301. doi:10.1163/156853968X00298.
- "Osservazioni comparativo ed esperimenti sulle modalita del corteg-giamento nelle cinque species europes del gruppo "obscura"". Cite journal requires
|journal=(help) - Spieth, Herman T. (1952). "Mating behavior within the genus Drosophila (Diptera)". Bulletin of the American Museum of Natural History. 99: 395–474. hdl:2246/934.
- "Steele RH. Courtship feeding in Drosophila subobscura. 2. Courtship feeding by males influences female mate choice, Animal Behavio ,1986, vol. 34, 1099-1108". Animal Behaviour. 34 (4): 1099–1108. August 1986. doi:10.1016/S0003-3472(86)80169-5. Retrieved 24 September 2019.
- "Steele RH. Courtship feeding in Drosophila subobscura. 1. The nutritional significance of courtship feeding, Animal Behavior, 1986, vol. 34, pg. 1087-1098". Retrieved 24 September 2019.
- Immonen, Elina; Hoikkala, Anneli; Kazem, Anahita J.N.; Ritchie, Michael G. (2009). "When are vomiting males attractive? Sexual selection on condition-dependent nuptial feeding in Drosophila subobscura". Behavioral Ecology. 20 (2): 289–295. doi:10.1093/beheco/arp008.
- Steele, Richard H.; Partridge, Linda (1988). "A courtship advantage for small males in Drosophila subobscura". Animal Behaviour. 36 (4): 1190–1197. doi:10.1016/S0003-3472(88)80078-2.
- Walsh, B. S.; Heys, C.; Lewis, Z. (2017). "Gut microbiota influences female choice and fecundity in the nuptial gift-giving species, Drosophila subobscura (Diptera: Drosophilidae)". European Journal of Entomology. 114: 439–445. doi:10.14411/eje.2017.056.
- "Emmens CW. The morphology of the nucleus in the salivary glands of four species of Drosophila (D. melanogaster, D. immigrans, D. funebris and D. subobscura). Z Zellforsch u mikroskop Anal. 1937;26:1–20". doi:10.1007/BF00368251. Cite journal requires
|journal=(help) - "Mainx F, Koske T, Smital E. Untersuchungen über dier chromosomale Struktur europaischer Vertreter der Drosophila obscura Gruppe. Z Indukt Abstamm Vererbungsl. 1953;85:354–72". Cite journal requires
|journal=(help) - Loukas, M.; Krimbas, C. B.; Mavragani-Tsipidou, P.; Kastritsis, c. D. (1979). "Genetics of Drosophila subobscura populations". Journal of Heredity. 70 (1): 17–26. doi:10.1093/oxfordjournals.jhered.a109181. PMID 112146.
- Vergini, Y.; Krimbas, C. B.; Loukas, M. (October 1979). "Loukas M, Krimbas CB, Vergini Y. The genetics of Drosophila subobscura populations. IX. Studies on linkage disequilibrium in four natural populations. Genetics. 1979;93:497–523". Genetics. 93 (2): 497–523. PMC 1214095. PMID 535729. Retrieved 24 September 2019.
- "Santos J, Serra L, Solé E, Pascual M. FISH mapping of microsatellite loci from Drosophila subobscura and its comparison to related species. Chromosom Res. 2010;18:213–26". 2010. CiteSeerX 10.1.1.709.5350. Cite journal requires
|journal=(help) - Powell JR. Progress and prospects in evolutionary biology: the Drosophila model. New York: Oxford University Press; 1997.
- Santos, M.; Iriarte, P. F.; Céspedes, W.; Balanyà, J.; Fontdevila, A.; Serra, L. (2004). "Swift laboratory thermal evolution of wing shape (but not size) in Drosophila subobscura and its relationship with chromosomal inversion polymorphism". Journal of Evolutionary Biology. 17 (4): 841–855. doi:10.1111/j.1420-9101.2004.00721.x. PMID 15271084. S2CID 16488003.
- Matos, Margarida; Simões, Pedro; Duarte, Ana; Rego, Carla; Avelar, Teresa; Rose, Michael R. (2004). "Convergence to a novel environment: comparative method versus experimental evolution". Evolution. 58 (7): 1503–1510. JSTOR 3449375. PMID 15341152.
- Simões, P.; Santos, J.; Fragata, I.; Mueller, L. D.; Rose, M. R.; Matos, M. (2008). "How repeatable is adaptive evolution? The role of geographical origin and founder effects in laboratory adaptation". Evolution. 62 (8): 1817–29. doi:10.1111/j.1558-5646.2008.00423.x. PMID 18489721.
- Zivanovic, G.; Mestres, F. (2011). "Changes in chromosomal polymorphism and global warming: The case of Drosophila subobscura from Apatin (Serbia)". Genetics and Molecular Biology. 34 (3): 489–495. doi:10.1590/S1415-47572011000300020. PMC 3168192. PMID 21931524.
- Dolgova, O.; Rego, C.; Calabria, G.; Balanyà, J.; Pascual, M.; Rezende, E. L.; Santos, M. (2010). "Genetic constraints for thermal coadaptation in Drosophila subobscura". BMC Evolutionary Biology. 10: 363. doi:10.1186/1471-2148-10-363. PMC 3003277. PMID 21108788.
- Krimbas, C.B. & Loukas,M.(1980) Inversion Polymorphism of Drosophila subobscura Evol.Biol.12,163-234.
- King, Robert (2012-12-06). Dobzhansky T, Powell JR: Drosophila pseudoobscura and its American relatives, D. persimilis and D. miranda. Hand Book of Genetics. Edited by: King RC. 1975, New York: Plenum, 3: 537-587. ISBN 9781461571483.
- Powell JR. Progress and Prospects in Evolutionary Biology: The Drosophila Model. 1997, New York: Oxford Univ Press.
- Powell JR & Krimbas, C.B., Drosophila Inversion Polymorphism / Edition 1.
- Balanya, J.; Oller, Josep M.; Huey, Raymond B.; Gilchrist, George W.; Serra, Luis (2006). "Global Genetic Change Tracks Global Climate Warming in Drosophila subobscura". Science. 313 (5794): 1773–1775. Bibcode:2006Sci...313.1773B. doi:10.1126/science.1131002. PMID 16946033.
External links
- Bioimages images of Drosophila subobscura