Danger model
The danger model is a theory of how the immune system works. It is based on the idea that the immune system does not distinguish between self and non-self, but rather between things that might cause damage and things that will not.
Brief history of immunologic models
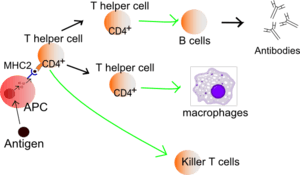
The first immunologic model was called the Self Non-self model and was suggested by Burnet and Frank Fenner in 1949,[1] and refined over the years.[2] It supposes that the immune system distinguishes between self, which is tolerated, and non-self, which is attacked and destroyed. According to this theory the chief cell of the immune system is the B lymphocyte, which is activated when it recognizes any non-self structure. Later it was discovered that in order to become activated, the B lymphocyte needs help from a CD4+ T helper cell, and that this T helper cell in turn also required a co-stimulatory signal from an antigen-presenting cell (APC) to be activated. The Self Non-self model developed according to these findings. Because APCs are not antigen-specific and are also capable of processing self structures, in 1989 Charles Janeway proposed a new theory, the so-called Infectious Non-self Model.[3] It is based on the idea that APCs are activated via pattern recognition receptors (PRRs) which recognize evolutionary distant conserved patterns. These pathogen-associated molecular patterns (PAMPs) on such organisms as bacteria are recognized as infectious non-self, whereas PRRs are not activated by non-infectious self. However, none of these models can explain non-cytopathic virus infections, graft rejection or anti-tumor immunity.[4]
The danger model
In 1994[5] a new immunologic model was suggested by Polly Matzinger. She suggested that the immune system does not distinguish between self and nonself, but discriminates between dangerous and safe by recognition of pathogens or alarm signals from injured or stressed cells and tissues.[6]
According to this theory, the most important for stimulation of immune response are normal tissues. When tissue cells are distressed because of injury, infection and so on, they start to secrete or express on their surface so called "Danger signals". "Danger signals" are also introduced into extracellular space when stressed cells die by immunologic not-silent cell death such as necrosis or pyroptosis (as opposed to apoptosis, controlled cell death). This model also suggests that, despite their potential immunogenicity, neoplastic tumors do not induce significant immune responses to induce the destruction of the malignant cells. According to the danger model, the immune surveillance system fails to detect tumor antigens because transformed cells do not send any danger signals which could activate dendritic cells and initiate an immune response. "Danger signals" are normal intracellular molecules that are not found in the extracellular space under physiological conditions. The danger model has evolved over the years.[7] "Danger signals" include DNA, RNA, heat shock proteins (Hsps), hyaluronic acid, serum amyloid A protein, ATP, uric acid and also cytokines like interferon-α, interleukin-1β, CD40L and so on.[4] [6][8]
The damage-associated molecular pattern (DAMP) model
The "danger model" suggests that, upon cellular necrosis, cryptic molecules in normal cells might be liberated from the cytoplasm into the tissue microenvironment, new molecules might be expressed on the surface of killed cells, or new molecules might be synthesized by cells undergoing necrosis. In comparison, the "DAMP model" [9] suggests that any nonfunctional molecules (regardless of their cellular location; extracellular or intracellular) that were denatured, oxidized, or had disintegrated quaternary structures might have unique biophysical patterns such as low water-solubility or stretches of polyanions on their surface.
Pattern recognition receptor for danger signal, DAMP and PAMP
"Danger signals" and DAMPs are together with pathogen-associated molecular patterns (PAMPs) called alarmins and they are recognized by pattern recognition receptors (PRRs) of APC cells. PRRs include Toll-like receptors (TLR), nucleotide oligomerization domain (NOD)-like receptors,[10] retinoic acid inducible gene-I (RIG-I)-like receptors[11] and C-type lectin-like receptors.[12] They are not only at the surface of these cells, but we can find them in cytoplasm and incorporated in the membrane of endolysosomes. Stimulation of PRRs leads to activation of APC cell to process antigen, upregulate expression of costimulatory molecules and present antigen to T helper cells.[6]
The danger model is a new perspective on adaptive and innate immunity. In the past innate immunity was suggested to be a minor part of the immune system — in contrast, adaptive immunity was thought to be the most important and effective part of the immune system. According to the danger model there is no adaptive immunity without the innate part. This is because APCs like dendritic cells are essential for activation of T lymphocytes and B lymphocytes, which after activation produce specific antibodies. In the case of dendritic cells deficiency, like in common variable immunodeficiency (CVID), patients suffer from hypogammaglobulinemia and from primary or secondary defects in T-cell functions.[4][13]
References
- Burnet FM; Fenner F (1949). The Production of Antibodies (2nd ed.). Melbourne: Macmillan.
- Burnet FM (1969). Cellular Immunology: Self and Notself. Cambridge: Cambridge University Press.
- Janeway, C. A. (1989-01-01). "Approaching the asymptote? Evolution and revolution in immunology". Cold Spring Harbor Symposia on Quantitative Biology. 54 (1): 1–13. doi:10.1101/sqb.1989.054.01.003. ISSN 0091-7451. PMID 2700931.
- Matzinger P (2002). "The danger model: a renewed sense of self". Science. 296 (5566): 301–5. CiteSeerX 10.1.1.127.558. doi:10.1126/science.1071059. PMID 11951032.
- Matzinger, P (2003-11-28). "Tolerance, Danger, and the Extended Family". Annual Review of Immunology. 12 (1): 991–1045. doi:10.1146/annurev.iy.12.040194.005015. PMID 8011301.
- Hallenbeck J, Del Zoppo G, Jacobs T, Hakim A, et al. (2006). "Immunomodulation strategies for preventing vascular disease of the brain and heart: workshop summary". Stroke. 37 (12): 3035–42. doi:10.1161/01.STR.0000248836.82538.ee. PMC 1853372. PMID 17082471.
- Pradeu, Thomas; Cooper, Edwin L. (2012-01-01). "The danger theory: 20 years later". Frontiers in Immunology. 3: 287. doi:10.3389/fimmu.2012.00287. ISSN 1664-3224. PMC 3443751. PMID 23060876.
- Jounai N, Kobiyama K, Takeshita F, Ishii KJ (2012). "Recognition of damage-associated molecular patterns related to nucleic acids during inflammation and vaccination". Front Cell Infect Microbiol. 2: 168. doi:10.3389/fcimb.2012.00168. PMC 3539075. PMID 23316484.
- Seong S, Matzinger P (2004). "Hydrophobicity: an ancient damage-associated molecular pattern that initiates innate immune responses". Nature Reviews Immunology. 4 (6): 469–478. doi:10.1038/nri1372. PMID 15173835.
- Tanti JF, Ceppo F, Jager J, Berthou F (2012). "Implication of inflammatory signaling pathways in obesity-induced insulin resistance". Front Endocrinol (Lausanne). 3: 181. doi:10.3389/fendo.2012.00181. PMC 3539134. PMID 23316186.
- Beckham SA, Brouwer J, Roth A, Wang D, et al. (2012). "Conformational rearrangements og RIG-I receptor on formation of a multiprotein: dsRNA assembly". Nucleic Acids Res. 41 (5): 3436–45. doi:10.1093/nar/gks1477. PMC 3597671. PMID 23325848.
- Kuroki K, Furukawa A, Maenaka K (2012). "Molecular recognition of paired receptors in the immune system". Front Microbiol. 3: 429. doi:10.3389/fmicb.2012.00429. PMC 3533184. PMID 23293633.
- Bayry J, Lacroix-Desmazes S, Kazatchkine MD, Galicier L, et al. (2004). "Common variable immunodeficiency is associated with defective functions of dendritic cells". Blood. 104 (8): 2441–3. doi:10.1182/blood-2004-04-1325. PMID 15226176.