Consumer demand tests (animals)
Consumer demand tests for animals are studies designed to measure the relative strength of an animal's motivation to obtain resources such as different food items. The test results are analogous to human patterns of purchasing resources with a limited income.[1][2] For humans, the cost of resources is usually measured in money; in animal studies the cost is usually represented by energy required, time taken or a risk of injury.[3] Costs of resources can be imposed on animals by an operant task (e.g. lever-pressing), a natural aversion (e.g. crossing water), or a homeostatic challenge (e.g. increased body temperature). Humans usually decrease the amount of an item purchased (or consumed) as the cost of that item increases. Similarly, animals tend to consume less of an item as the cost of that item increases (e.g. more lever presses required).[4][5] Such demand tests quantify the strength of motivation animals have for resources whilst avoiding anthropomorphism and anthropocentrism.[6]
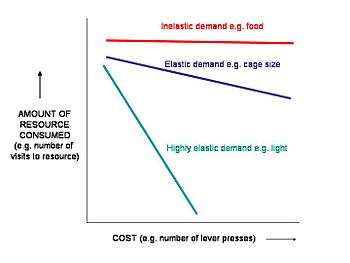
Using consumer demand tests one can empirically determine the strength of motivation animals have for a definite need (e.g. food, water) and also for resources we humans might perceive as a luxury or unnecessary but animals might not (e.g. sand for dustbathing[7] or additional space for caged mice[8]). By comparing the strength of motivation for the resource with that for a definite need, we can measure the importance of a resource as perceived by the animals. Animals will be most highly motivated to interact with resources they absolutely need, highly motivated for resources that they perceive as most improving their welfare, and less motivated for resources they perceive as less important. Furthermore, Argument by analogy indicates that as with humans, it is more likely that animals will experience negative affective states (e.g. frustration, anxiety) if they are not provided with the resources for which they show high motivation.[3]
Various other aspects of the animal's behaviour can be measured to aid understanding of motivation for resources, e.g. latency (delay) to approach the point of access, speed of incurring the cost, time with each resource, or the range of activities with each of the resources. These measures can be recorded either by the experimenter or by motion detecting software. Prior to testing, the animals are usually given the opportunity to explore the apparatus and variants to habituate and reduce the effects of novelty.
Terminology
The rate (i.e. regression line) at which the animal decreases its acquisition or consumption of a resource as the cost increases is known as the elasticity of demand. A steep slope of decreasing access indicates a relatively low motivation for a resource, sometimes called 'high elasticity'; a shallow slope indicates relatively high motivation for a resource, sometimes called 'low elasticity', or 'inelastic demand.'
The 'break point' is the cost at which inelastic demand becomes elastic, i.e. the cost at which constant consumption begins to decrease.
In human economics and consumer theory, a Giffen good is a resource which is paradoxically consumed more as the cost rises, violating the law of demand. In normal situations, as the price of a resource increases, the substitution effect causes consumers to purchase less of it and more of substitute goods. In the Giffen good situation, the income effect dominates, leading people to buy more of the good, even as its price rises.
Types of cost
Homeostatic challenge
- Body temperature[16]
Examples
Flooring
Manser et al.[10] showed that laboratory rats were motivated to lift a door weighing 83% of their body weight to allow them to rest on a solid floor rather than on a grid floor, despite their having been kept on grid floors for over 6 months.
Lighting
Baldwin showed that when animals were given control of their lighting with the equivalent of an on/off switch, pigs kept lights on for 72% of the time and sheep for 82%. However, when the pigs had to work for the light by keeping their snout within a photo-beam, they only kept the lights on for 0.5% of the time, indicating that light was a weak reinforcement for this species.[12] Savory and Duncan showed that individual hens kept in a background of darkness were prepared to work for 4 hours of light per day.[17]
Burrowing substrate
Sherwin et al. examined the strength of motivation for burrowing substrate in laboratory mice. Despite an increasing cost of gaining access, the mice continued to work to visit the burrowing substrate.[18] In addition, it was shown that it was the performance of burrowing behaviour that was important to the mice, not simply the functional consequences of the behaviour. King and Welsman showed that when bar pressing gave deermice access to sand, they increased their rate of bar pressing as the number of presses to access the sand was increased.[19]
Nest box
Duncan and Kite showed that hens were highly motivated to gain access to a nest box, particularly immediately prior to oviposition. The hens would push a weighted door, or walk through water or an air blast to reach a nest box. Duncan and Kite suggested the strength of this motivation was equivalent to that of the strength of motivation to feed after 20 hours deprivation.[20]
Social contact
Several studies have examined the motivation of animals for social contact either with their offspring or conspecifics.[21][22][23]
See also
- Preference tests (animals)
- Consumer theory
References
- Lea, S.E.G., (1978). The psychology and economics of demand. Psychological Bulletin, 85:441–466
- Dawkins, M.S., (1983). Battery hens name their price: consumer demand theory and the measurement of ethological "needs". Animal Behaviour, 31: 1195–1205
- Dawkins, M.S., (1990). From an animal's point of view: motivation, fitness, and animal welfare. Behavioral and Brain Sciences, 13: 1–61
- Duncan, I.J.H., (1992). Measuring preferences and the strength of preferences. Poultry Science, 71: 658–663
- Sherwin, C.M., (1996). Laboratory mice persist in gaining access to resources: a method of assessing the importance of environmental features. Applied Animal Behaviour Science, 48: 203–214
- "Sherwin, C.M. Validating refinements to laboratory housing:asking the animals". Retrieved September 11, 2012.
- Faure, J.M. and Lagadic, H., (1994). Elasticity of demand for food and sand in laying hens subjected to variable wind speed. Applied Animal Behaviour Science, 42: 49–59
- Sherwin, C.M. and Nicol, C.J., (1997). Behavioural demand functions of caged laboratory mice for additional space. Animal Behaviour, 53: 67–74
- Sherwin, C.M., (1998). The use and perceived importance of three resources which provide caged laboratory mice with the opportunity for extended locomotion. Applied Animal Behaviour Science, 55: 353–367
- Manser, C.E., Elliott, H., Morris, T.H. and Broom, D,M., (1996). The use of a novel operant test to determine the strength of preference for flooring in laboratory rats. Laboratory Animals, 30: 1–6
- Olsson, I.A.S. and Keeling, L.J., (2002). The push-door for measuring motivation in hens: laying hens are motivated to perch at night. Animal Welfare, 11: 11–19
- Baldwin, B.A. (1979). Operant studies on the behaviour of pigs and sheep in relation to the physical environment. Journal of Animal Science, 49: 1125-1134
- Collier,. G.H., Johnson, D.F., CyBulski, K.A. and McHale, C.A., (1990). Activity patterns in rats (Rattus norvegicus) as a function of the cost of access to four resources. Journal od Comparative Psychology, 104: 53-65
- Sherwin, C.M. and Nicol, C.J., (1996). Reorganisation of behaviour in laboratory mice, Mus musculus, with varying cost of access to resources. Animal Behaviour, 51: 1087–1093
- Guerra, R.F. and Ades, C. (2002). An analysis of travel costs on transport of load and nest building in golden hamster. Behaviour Proceedings, 57: 7-28
- Johnson, K.G. and Cabanac, M., (1982). Homeostatic competition between food intake and temperature regulation in rats. Physiology & Behavior, 28: 675–679
- Savory, C.J. and Duncan, I.J.H. (1982.) Voluntary regulation of lighting by domestic fowls in skinner boxes. Applied Animal Ethology, 9: 73-81
- Sherwin, C.M., Haug, E., Terkelsen, N. and Vadgama, M., (2004). Studies on the motivation for burrowing by laboratory mice. Applied Animal Behaviour Science, 88: 343-358
- King, J.A. and Weisman, R.G., (1964) Sand digging contingent upon bar pressing in deermice (Peomyscus). Animal Behaviour, 12: 446-450
- Duncan, I.J.H. and Kite, V.G., (1987) Some investigations into motivation in the domestic fowl. Applied Animal Behaviour Science, 18: 387-388
- Wilsoncroft, W.E. (1969). Babies by bar-press: maternal behavior in the rat. Behavior Research Methods and Instrumentation, 1: 229-230
- Van Hemel, S.B. (1973). Pup retrieving as a reinforcer in nulliparous mice. Journal of Experimental Analysis of Behaviour, 19: 233-238
- Mills, A.D. and Faure, J.M., (1990). The treadmill test for the measurement of social motivation in Phasianidae chicks. Medical Science Research 18: 179-180