Alpha-keratin
Alpha-keratin, or α-keratin, is a type of keratin found in vertebrates. This protein is the primary component in hairs, horns, mammalian claws, nails and the epidermis layer of the skin. α-keratin is a fibrous structural protein, meaning it is made up of amino acids that form a repeating secondary structure. The secondary structure of α-keratin is very similar to that of a traditional protein α-helix and forms a coiled coil.[1] Due to its tightly wound structure, it can function as one of the strongest biological materials and has various functions in mammals, from predatory claws to hair for warmth. α-keratin is synthesized through protein biosynthesis, utilizing transcription and translation, but as the cell matures and is full of α-keratin, it dies, creating a strong non-vascular unit of keratinized tissue.[2]
Structure
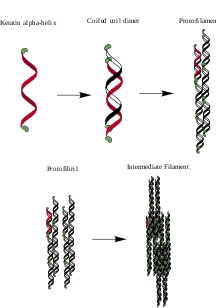
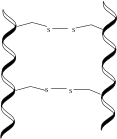
α-keratin is a polypeptide chain, typically high in alanine, leucine, arginine, and cysteine, that forms a right-handed α-helix.[3][4] Two of these polypeptide chains twist together to form a left-handed helical structure known as a coiled coil. These coiled coil dimers, approximately 45 nm long, are bonded together with disulfide bonds, utilizing the many cysteine amino acids found in α-keratins.[2] The dimers then align, their termini bonding with the termini of other dimers, and two of these new chains bond length-wise, all through disulfide bonds, to form a protofilament.[5] Two protofilaments aggregate to form a protofibril, and four protofibrils polymerize to form the intermediate filament (IF). The IF is the basic subunit of α-keratins. These IFs are able to condense into a super-coil formation of about 7 nm in diameter, and can be type I, acidic, or type II, basic. The IFs are finally embedded in a keratin matrix that either is high in cysteine or glycine, tyrosine, and phenylalanine residues. The different types, alignments, and matrices of these IFs account for the large variation in α-keratin structures found in mammals.[6]
Biochemistry
Synthesis
α-keratin synthesis begins near focal adhesions on the cell membrane. There, the keratin filament precursors go through a process known as nucleation, where the keratin precursors of dimers and filaments elongate, fuse, and bundle together.[2] As this synthesis is occurring, the keratin filament precursors are transported by actin fibers in the cell towards the nucleus. There, the alpha-keratin intermediate filaments will collect and form networks of structure dictated by the use of the keratin cell as the nucleus simultaneously degrades.[7] However, if necessary, instead of continuing to grow, the keratin complex will disassemble into non-filamentous keratin precursors that can diffuse throughout the cell cytoplasm. These keratin filaments will be able to be used in future keratin synthesis, either to re-organize the final structure or create a different keratin complex. When the cell has been filled with the correct keratin and structured correctly, it undergoes keratin stabilization and dies, a form of programmed cell death. This results in a fully matured, non-vascular keratin cell.[8] These fully matured, or cornified, alpha-keratin cells are the main components of hair, the outer layer of nails and horns, and the epidermis layer of the skin.[9]
Properties
The property of most biological importance of alpha-keratin is its structural stability. When exposed to mechanical stress, α-keratin structures can retain their shape and therefore can protect what they surround.[10] Under high tension, alpha-keratin can even change into beta-keratin, a stronger keratin formation that has a secondary structure of beta-pleated sheets.[11] Alpha-keratin tissues also show signs of viscoelasticity, allowing them to both be able to stretch and absorb impact to a degree, though they are not impervious to fracture. Alpha-keratin strength is also affected by water content in the intermediate filament matrix; higher water content decreases the strength and stiffness of the keratin cell due to their effect on the various hydrogen bonds in the alpha-keratin network.[2]
Characterization
Type I and type II
Alpha-keratins proteins can be one of two types: type I or type II. There are 54 keratin genes in humans, 28 of which code for type I, and 26 for type II.[12] Type I proteins are acidic, meaning they contain more acidic amino acids, such as aspartic acid, while type II proteins are basic, meaning they contain more basic amino acids, such as lysine.[13] This differentiation is especially important in alpha-keratins because in the synthesis of its sub-unit dimer, the coiled coil, one protein coil must be type I, while the other must be type II.[2] Even within type I and II, there are acidic and basic keratins that are particularly complementary within each organism. For example, in human skin, K5, a type II alpha keratin, pairs primarily with K14, a type I alpha-keratin, to form the alpha-keratin complex of the epidermis layer of cells in the skin.[14]
Hard and soft
Hard alpha-keratins, such as those found in nails, have a higher cysteine content in their primary structure. This causes an increase in disulfide bonds that are able to stabilize the keratin structure, allowing it to resist a higher level of force before fracture. On the other hand, soft alpha-keratins, such as ones found in the skin, contain a comparatively smaller amount of disulfide bonds, making their structure more flexible.[1]
References
- G., Voet, Judith; W., Pratt, Charlotte (2016-02-29). Fundamentals of biochemistry : life at the molecular level. ISBN 9781118918401. OCLC 910538334.
- Wang, Bin; Yang, Wen; McKittrick, Joanna; Meyers, Marc André (2016-03-01). "Keratin: Structure, mechanical properties, occurrence in biological organisms, and efforts at bioinspiration" (PDF). Progress in Materials Science. 76: 229–318. doi:10.1016/j.pmatsci.2015.06.001.
- Burkhard, Peter; Stetefeld, Jörg; Strelkov, Sergei V (2001). "Coiled coils: a highly versatile protein folding motif". Trends in Cell Biology. 11 (2): 82–88. doi:10.1016/s0962-8924(00)01898-5. PMID 11166216.
- Pace, C N; Scholtz, J M (1998-07-01). "A helix propensity scale based on experimental studies of peptides and proteins". Biophysical Journal. 75 (1): 422–427. doi:10.1016/S0006-3495(98)77529-0. ISSN 0006-3495. PMC 1299714. PMID 9649402.
- Steinert, Peter M.; Steven, Alasdair C.; Roop, Dennis R. (1985). "The molecular biology of intermediate filaments". Cell. 42 (2): 411–419. doi:10.1016/0092-8674(85)90098-4. PMID 2411418.
- McKittrick, J.; Chen, P.-Y.; Bodde, S. G.; Yang, W.; Novitskaya, E. E.; Meyers, M. A. (2012-04-03). "The Structure, Functions, and Mechanical Properties of Keratin". JOM. 64 (4): 449–468. doi:10.1007/s11837-012-0302-8. ISSN 1047-4838.
- Windoffer, Reinhard; Beil, Michael; Magin, Thomas M.; Leube, Rudolf E. (2011-09-05). "Cytoskeleton in motion: the dynamics of keratin intermediate filaments in epithelia". The Journal of Cell Biology. 194 (5): 669–678. doi:10.1083/jcb.201008095. ISSN 0021-9525. PMC 3171125. PMID 21893596.
- Kölsch, Anne; Windoffer, Reinhard; Würflinger, Thomas; Aach, Til; Leube, Rudolf E. (2010-07-01). "The keratin-filament cycle of assembly and disassembly". J Cell Sci. 123 (13): 2266–2272. doi:10.1242/jcs.068080. ISSN 0021-9533. PMID 20554896.
- Eckhart, Leopold; Lippens, Saskia; Tschachler, Erwin; Declercq, Wim (2013-12-01). "Cell death by cornification". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1833 (12): 3471–3480. doi:10.1016/j.bbamcr.2013.06.010. PMID 23792051.
- Pan, Xiaoou; Hobbs, Ryan P; Coulombe, Pierre A (2013). "The expanding significance of keratin intermediate filaments in normal and diseased epithelia". Current Opinion in Cell Biology. 25 (1): 47–56. doi:10.1016/j.ceb.2012.10.018. PMC 3578078. PMID 23270662.
- Kreplak, L.; Doucet, J.; Dumas, P.; Briki, F. (2004-07-01). "New Aspects of the α-Helix to β-Sheet Transition in Stretched Hard α-Keratin Fibers". Biophysical Journal. 87 (1): 640–647. doi:10.1529/biophysj.103.036749. PMC 1304386. PMID 15240497.
- Moll, Roland; Divo, Markus; Langbein, Lutz (2017-03-07). "The human keratins: biology and pathology". Histochemistry and Cell Biology. 129 (6): 705–733. doi:10.1007/s00418-008-0435-6. ISSN 0948-6143. PMC 2386534. PMID 18461349.
- Strnad, Pavel; Usachov, Valentyn; Debes, Cedric; Gräter, Frauke; Parry, David A. D.; Omary, M. Bishr (2011-12-15). "Unique amino acid signatures that are evolutionarily conserved distinguish simple-type, epidermal and hair keratins". Journal of Cell Science. 124 (24): 4221–4232. doi:10.1242/jcs.089516. ISSN 0021-9533. PMC 3258107. PMID 22215855.
- Lee, Chang-Hun; Coulombe, Pierre A. (2009-08-10). "Self-organization of keratin intermediate filaments into cross-linked networks". The Journal of Cell Biology. 186 (3): 409–421. doi:10.1083/jcb.200810196. ISSN 0021-9525. PMC 2728393. PMID 19651890.